disAMR Hosts VIP Performance of Lifeline at Southwark Playhouse Elephant
The Southwark Playhouse Elephant in London became the setting for something rare, a moment where science, medicine, policy, and the arts converged around one of the most urgent challenges facing modern medicine. disAMR hosted a VIP performance of Lifeline, the acclaimed musical that has become one of the most powerful and unconventional voices in the global fight against antimicrobial resistance (AMR).
The evening brought together life sciences leaders, policy makers, investors, and advocates, some deeply embedded in the AMR community, others in conversation with disAMR exploring the best ways to contribute, for a performance that combined professional West End theatre with something no conventional production can offer: a chorus of real-life doctors, nurses, scientists, microbiologists, and health workers who at the end of the performance shared their own stories from the frontlines of the resistance crisis.

“Lifeline reflects disAMR’s belief that human stories are needed to move people to action, and the importance of making the AMR crisis impossible to look away from because it can affect anyone,” said Mark Bamforth OBE, CEO of disAMR. “This evening was about bringing together the people who are fighting this battle every day and those outside of the traditional AMR ecosystem, reminding and educating why this work matters.”
Lifeline is the work of composer and lyricist Robin Hiley and book writer Becky Hope-Palmer, produced by The Charades Theatre Company. Its origins trace back to 2016, when infectious disease clinician Meghan Perry approached Hiley with an idea: to bring the story of antibiotic resistance to audiences who might never encounter it through a journal article or policy briefing. What began as a children’s play about Alexander Fleming grew, over nearly a decade, into a full-scale musical that has sold out runs at the Edinburgh Fringe Festival, achieved off-Broadway success at New York’s Pershing Square Signature Center, and made history in 2024 as the first musical ever performed on the floor of the United Nations, closing the Global High-Level Meeting on AMR to a standing ovation from diplomats representing nearly 200 countries.
The production weaves together two narratives across time. In 1950, Alexander Fleming, at the height of his fame following the discovery of penicillin, fell in love with Dr. Amalia Koutsouri-Voureka, a member of his research team and a Greek freedom fighter. In present-day Edinburgh, junior doctor Jess faces the unthinkable when her estranged childhood sweetheart is admitted to her hospital with a drug-resistant infection. The two stories mirror each other over seven decades, connecting Fleming’s prophetic Nobel Prize warning about the misuse of antibiotics to the reality he predicted. The result is a score that moves between Scottish folk-infused foot-stomping rhythms and moments of profound emotional weight, serving as a reminder that behind every AMR statistic is a human story.
For disAMR, sponsoring Lifeline is one expression of the organization’s mission. Building on the foundation of AMR Insights, which has served the global AMR community for eight years, disAMR is working to consider the AMR crisis from a different perspective and marshal resources and stakeholders not previously associated with efforts to diminish AMR.
“Annually, AMR is projected to contribute to 10 million deaths by 2050, and the world still doesn’t fully appreciate the scale of what is at stake,” said Richard Snyder, PhD, Chief Scientific Officer of disAMR. “Science alone will not solve this. We need awareness, advocacy, and new thinking. Lifeline is helping by bringing biology, economics, and the human cost of AMR to the forefront.”
The evening began with welcome drinks and the kind of cross-sector conversation that disAMR seeks to enable: researchers alongside investors, clinicians alongside communicators, policy leaders alongside advocates. Following the performance, guests gathered for dessert and continued dialogue.
“What struck me most was the energy in the room after the final curtain,” said Nigel Walker, COO of disAMR. “Lifeline is moving people by reminding us that AMR is not just a scientific or economic problem. It is a human one.”
Through engaging and creative storytelling, Lifeline is bringing new voices and talents to the table to help advance the fight against AMR. Similarly, disAMR’s central focus is to examine the AMR crisis from new perspectives and to bring more resources to the table. If joining us in this effort speaks to you, please contact us.
When most people think about antimicrobial resistance (AMR), they picture bacteria, the superbugs that have dominated headlines, policy discussions, and research funding for decades. But quietly, and largely out of public view, a parallel AMR crisis is unfolding in the fungal world. Drug-resistant fungi are spreading, killing millions, and receiving a fraction of the scientific and policy attention the threat demands.
Paul Verweij, professor of clinical mycology at Radboud University Medical Center in the Netherlands and one of the world’s foremost experts on fungal disease, describes what is happening as a “silent surge.” It is an appropriate phrase, as approximately 3.8 million people die each year from fungal infections, a number that has nearly doubled in the past decade, yet antifungal resistance is often not front and center in the AMR conversations that shape global health policy and research investment.
Wide Range of Fungal Infection Severity
Fungal infections span an enormous range of severity. At one end of the spectrum are common, manageable conditions like athlete’s foot, dandruff, toenail infections, and vaginal yeast infections. These conditions affect millions of people annually and are typically treatable with standard antifungal medications. At the other end are life-threatening infections that strike the most vulnerable: people undergoing chemotherapy, organ transplant recipients, patients in intensive care, and anyone with a compromised immune system.
Candida auris, a yeast first identified at a Tokyo hospital in 2009, has since spread to health facilities worldwide, exemplifying the threat it poses. Resistant to multiple classes of antifungal drugs and difficult to eliminate from hospital environments, C. auris kills between 30% and 60% of the people it infects. In low- and middle-income countries, where diagnostic capacity is limited and infection control resources are stretched, patients may die without the cause even being diagnosed.
Why Fungi Are So Difficult to Treat
Part of the reason antifungal resistance has been underestimated is that fungi present uniquely difficult scientific challenges. Unlike bacteria and viruses, fungal cells are structurally similar to human cells. This similarity means that developing drugs capable of killing fungi without damaging the patient is inherently complex. Specifically, the therapeutic window is narrower, and the risk of toxicity is higher. Lessons learned from bacterial AMR do not always translate directly to fungal resistance, which often operates through distinct molecular mechanisms.
Only four major classes of antifungal drugs are currently available — azoles, polyenes, allylamines, and the newer echinocandins — compared with the much broader antibiotic armamentarium for bacteria. As resistance to azoles, the most widely used class, continues to grow, the treatment options for serious fungal infections are narrowing rapidly. According to recent WHO analyses, only three drugs targeting the fungi that pose the greatest threat to human health are in late-stage clinical development.
Agriculture’s Contribution to Antifungal Resistance
One of the most concerning dimensions of antifungal resistance is its agricultural origin. Azole fungicides, chemically related to the azole drugs used to treat human fungal infections, are used extensively to protect crops from fungal disease. Without them, agricultural yield losses could reach 30 to 40 percent. But as environmental fungi develop resistance to agricultural fungicides, they also develop resistance to closely related drugs used in human health. Aspergillus fumigatus, a common mold that can cause severe lung infections in immunocompromised patients, is increasingly resistant to azoles, a pattern linked directly to fungicide exposure in agricultural environments.
This dynamic creates a genuine One Health dilemma, as the same chemical compounds that protect global food security are undermining the antifungal medicines that protect vulnerable patients. Some regulatory action is underway. For instance, the US Environmental Protection Agency has proposed factoring antifungal resistance risks into its assessment of new fungicides, but progress remains slow relative to the scale of the problem.
The Case for Urgent Action
In April 2025, Verweij and 50 co-authors published a call to action in Nature Medicine, urging the explicit incorporation of drug-resistant fungal disease into the 2026 update to the Global Action Plan on Antimicrobial Resistance.
Addressing antifungal resistance will require investment in basic research, the development of new diagnostic tools capable of rapidly identifying resistant strains, expanded clinical trial networks, and cross-sector collaboration that brings agricultural regulators into conversation with public health systems. While none of these efforts are simple undertakings, the global health community can no longer sideline this threat.
Why AMR Is Bigger Than a Public Health Problem
Antimicrobial resistance (AMR) is widely recognized as one of the most serious threats to global health. The World Health Organization (WHO) identifies AMR among the top 10 public health challenges facing humanity, owing to both its current burden and the trajectory of its spread.1 This designation places AMR alongside conditions that receive far greater public attention, even though its impact is often less visible and more diffuse.
In 2019, bacterial AMR was associated with an estimated 4.95 million deaths globally, including 1.27 million deaths directly attributable to resistant infections.2 These figures demonstrate that AMR is not merely a concern for the future but a growing contributor to global mortality in the present. As resistance continues to erode the effectiveness of existing therapies, formerly routine infections are becoming more complex, more resource-intensive, and in some cases, untreatable.
Viewing AMR strictly as a clinical or epidemiological issue risks underestimating its danger. Resistant infections do not remain confined to individual patients; they influence how care is delivered, how hospitals allocate resources, and how health systems maintain capacity under strain. The consequences extend well beyond the failure of an individual treatment: longer hospital stays, increased use of intensive care, and higher demand on resources.
For this reason, AMR is more accurately understood as a system-level risk. Its effects reverberate through healthcare delivery, economic productivity, and the broader resilience of society as a whole. Recognizing this shift in perspective is essential. AMR is a structural vulnerability that affects multiple interconnected systems and must be considered alongside other infrastructure risks.
The Hidden Dependency: Modern Medicine Runs on Antibiotics
Modern medicine relies on the premise that antimicrobial therapies will remain effective. Antibiotics do far more than treat infections; they are central to safety across a wide range of routine and advanced medical interventions. Procedures as varied as major surgery, cancer chemotherapy, organ transplantation, and dialysis all rely on the ability to prevent and control infections that arise when natural barriers are breached or immune systems are compromised.
This dependency is structural rather than incidental. Surgical procedures, even when performed under sterile conditions, carry a risk of infection that must be managed prophylactically and therapeutically. Chemotherapy suppresses immune function, which increases susceptibility to opportunistic infections. Transplant recipients require long-term immunosuppression, further elevating that risk. Dialysis patients undergo repeated vascular access and face persistent exposure to potential pathogens. In each case, antibiotics are critical to the viability of these interventions at scale.
As the effectiveness of antibiotics decreases, that protection weakens. Infections become harder to prevent and more difficult to treat, increasing the likelihood of complications, prolonged recovery, or treatment failure. Procedures that are now considered routine begin to carry higher levels of uncertainty and, in some cases, may no longer be feasible within acceptable risk thresholds.
This dynamic shifts the implications of AMR beyond the management of infectious diseases. Undermining the conditions required for their safe and reliable execution places core components of modern healthcare delivery at risk. In this context, antibiotics must be seen as enabling infrastructure for the entire medical system. As resistance spreads, not only are individual therapies compromised, but the operational foundation on which contemporary healthcare depends is as well.
Healthcare Systems Under Strain: Capacity, Not Just Cost
The impact of AMR on healthcare systems is often discussed in terms of cost, but its effects on capacity are just as consequential. Resistant infections are associated with longer hospital stays, greater reliance on intensive care, and increased use of specialized resources, all of which place sustained pressure on systems that are already highly constrained.3,4 These demands are not evenly distributed; they concentrate in high-acuity settings where capacity is limited and difficult to expand quickly.
Increasing resource intensity carries the risk of operational disruption. Outbreaks of resistant pathogens can require hospital ward closures and isolation procedures, diverting staff and equipment and interrupting routine care delivery. Procedures must be delayed or cancelled to contain transmission or to redirect capacity to more urgent needs. These disruptions ripple outward, affecting not only patients with resistant infections but also those awaiting elective or time-sensitive care.
The COVID-19 pandemic provided a clear illustration of how system stress can amplify these dynamics. During the first year of the pandemic, the United States saw a significant increase in antimicrobial-resistant infections, including tens of thousands of deaths associated with healthcare-associated infections.5 Under conditions of strain, infection control practices can be harder to maintain, antibiotic use may increase, and surveillance systems may be disrupted, creating an environment in which resistance can spread more readily.
AMR does more than increase the burden on healthcare systems; it reduces their effective capacity to deliver care. Beds remain occupied longer, staff and resources are diverted, and throughput declines. This pattern resembles infrastructure failure, where systems continue to operate but at diminished efficiency and reliability. As resistance grows, the gap between nominal capacity and functional capacity widens, with direct implications for access, quality, and resilience.
Economic Productivity and System-Wide Cost Escalation
The economic consequences of AMR extend beyond healthcare budgets. At a macroeconomic level, AMR has the potential to slow growth, disrupt labor markets, and exacerbate inequality. World Bank modeling suggests that under high-resistance scenarios, AMR could reduce global gross domestic product by up to 3.8% annually by 2050, with an estimated 28 million additional people pushed into poverty.6 These projections reflect not only the direct costs of treatment but also the broader effects of illness, reduced labor participation, and diminished productivity.
Within healthcare systems, the financial burden can already be measured. In the United States alone, antimicrobial-resistant infections are estimated to cost billions of dollars each year, driven by longer hospital stays, more intensive treatment requirements, and the need for more complex and expensive therapies.7,8 Earlier analyses have also linked AMR to tens of billions of dollars in combined direct healthcare costs and lost productivity, as well as millions of additional hospital days annually.3 These costs are not isolated; they accumulate across payers, providers, and public health systems and compound existing financial pressures.
At the workforce level, AMR contributes to lost working days, reduced labor force participation, and reduced working life expectancy. Resistant infections can prolong illness, delay recovery, and increase the likelihood of complications, all of which affect an individual’s ability to return to work or maintain productivity over time.9 These effects translate into measurable declines in labor output, particularly in scenarios where resistance becomes more widespread.

AMR as a National and Global Security Issue
AMR can no longer be viewed solely through a public health or even an economic lens. Governments increasingly treat it as a component of national and global security, reflecting its potential to disrupt critical systems and undermine preparedness. In the United States, AMR is incorporated into broader health security strategies, with federal agencies emphasizing coordinated, cross-sector responses to mitigate its impact.10 This reflects an understanding that infectious disease threats, including resistant pathogens, can affect not only population health but also institutional stability and crisis response capabilities.
The U.S. Department of Defense has taken this view further, identifying AMR as a factor that can overwhelm response capacity and generate cascading health, economic, and societal consequences.11 Within military contexts, the implications are direct. Resistant infections complicate the treatment of combat-related injuries, increasing the risk of severe outcomes, such as limb loss or death, and can compromise recovery timelines and operational readiness. Ongoing investment in infectious disease research, including AMR, reflects its relevance to maintaining force health and mission effectiveness.
At the international level, AMR has emerged as a priority for multilateral coordination. In 2024, United Nations member states adopted a political declaration committing to intensified action on AMR, including efforts to reduce the global burden of resistant infections.12 This consensus indicates that AMR is not only a technical or clinical challenge but a shared strategic concern with implications for global stability.
This convergence of public health, defense, and international policy perspectives underscores a broader shift in how AMR is understood. Governments are already treating it as a security issue because of its ability to degrade system performance, strain response capacity, and amplify the impact of other crises. While it may not always be explicitly labeled as an infrastructure risk, its effects closely align with those of other threats that compromise the reliability and resilience of essential systems.
Reframing AMR: A System-Level Infrastructure Risk
The evidence shows a pattern of AMR impacts that span multiple systems beyond its clinical and epidemiological dimensions. AMR increases the cost of care, reduces the effective capacity of healthcare delivery, disrupts routine operations, diminishes workforce productivity, and carries implications for national and global security.
These effects do not occur in isolation. Longer hospital stays and greater resource intensity constrain bed availability and staff capacity. Operational disruptions, such as ward closures or delayed procedures, reduce throughput and limit access to care. At the same time, increased illness and prolonged recovery reduce labor force participation and productivity, thereby reinforcing the economic burden. Security concerns add another layer, as resistant infections complicate emergency response and strain institutional readiness.
These dynamics resemble the failure modes observed in other forms of infrastructure degradation. Systems continue to function, but with reduced efficiency, diminished reliability, and less resilience to external shocks. The gap between nominal and functional capacity widens, making the system more vulnerable to disruption under stress.
Viewing AMR as an infrastructure risk captures this interconnected pattern. Antibiotics serve as enabling components of modern healthcare and, by extension, of broader economic and social systems. As their effectiveness declines, the systems that depend on them lose stability and performance. Recognizing AMR in these terms clarifies why its impact extends far beyond infection control and why addressing it requires coordinated action across healthcare, economic policy, and security domains.
Protecting Antibiotics Means Protecting System Capacity
Antibiotics are not simply another class of therapeutics. They enable the safe delivery of modern medicine and support the reliable functioning of healthcare systems, economies, and institutions. As AMR advances, that enabling role comes under increasing strain.
These pressures do not remain contained. They accumulate across systems, reducing efficiency in routine conditions and limiting the ability to respond to disruption. What is at risk is not only the effectiveness of individual treatments but the capacity of interconnected systems to operate as intended.
Efforts to address AMR must evolve beyond a narrow focus on stewardship and drug development alone. They require coordinated action across healthcare delivery, public health infrastructure, economic policy, and security planning.
A central challenge in addressing antimicrobial resistance is the misalignment between the nature of the risk and the structure of responses. The benefits of preserving antibiotic effectiveness manifest at the system level over long time horizons, but incentives are often defined at the level of individual institutions, products, or short-term policy cycles. Pharmaceutical innovation is constrained by limited commercial returns for antibiotics that must be used sparingly, while healthcare providers face immediate clinical and financial pressures that favor broader, precautionary use. At the policy level, interventions must be considered within electoral and budgetary cycles that align poorly with the slow progression of resistance. At the same time, responsibility for AMR is distributed across healthcare, agriculture, public health, and environmental systems, with no single entity accountable for managing its full impact.
This fragmentation complicates coordination and limits the ability to implement sustained, system-wide strategies. Current approaches tend to address individual components of the problem rather than the underlying conditions that allow resistance to accumulate, limiting their effectiveness in preserving antibiotic function as a shared infrastructure resource. They require coordinated action across healthcare delivery, public health infrastructure, economic policy, and security planning. Antibiotic effectiveness must be treated as a shared, finite resource that underpins critical system performance.
Protecting that resource is therefore a matter of system preservation. Maintaining antibiotic effectiveness means sustaining healthcare capacity, supporting economic productivity, and ensuring resilience in the face of future shocks. Recognizing AMR in these terms clarifies both the scale of the challenge and the urgency of response.
The Evolutionary Problem of Antimicrobial Resistance
Antimicrobial resistance (AMR) is typically framed as a clinical or pharmaceutical challenge, but at its core, it is an evolutionary process. Antibiotics create strong selective pressures within microbial populations. Bacteria that have acquired genetic changes (through one of several mechanisms) allowing them to survive drug exposure gain a temporary advantage, while susceptible strains are eliminated. Over time, this selection reshapes microbial communities, increasing the prevalence of resistant organisms.
These evolutionary dynamics can operate on fast time scales because bacterial populations are large and can undergo multiple rounds of replication rapidly. Random mutations, gene acquisition events, and recombination generate genetic variation continuously. When antibiotics are present, variants capable of surviving treatment can expand, allowing resistance traits to spread through bacterial populations. In this sense, the emergence of resistance is not an unusual biological event but a predictable outcome of sustained selective pressure.1
Viewing resistance through an evolutionary lens highlights a nuance familiar in evolutionary biology but less emphasized in the clinical context: resistance is not a binary property. The genetic changes that allow bacteria to evade antibiotics often alter other aspects of bacterial physiology. Those alterations can influence how well resistant strains compete with susceptible strains in different environments. As a result, the evolutionary consequences of resistance extend beyond survival in the presence of antibiotics and can continue to shape bacterial population dynamics after antibiotic pressure changes.
Viewing antimicrobial resistance as an evolutionary system suggests a different approach to stewardship: rather than simply reducing antibiotic use or rotating drugs on fixed schedules, clinicians may eventually be able to guide resistance trajectories using insights from evolutionary biology.
The Mechanisms That Create Resistance
Antibiotics exert their effects by disrupting essential bacterial processes, such as cell wall synthesis, protein production, DNA replication, or metabolism. Resistance emerges when bacteria acquire genetic changes that allow them to avoid or neutralize those effects. These changes may arise through spontaneous mutation or through horizontal gene transfer (HGT) from other microorganisms. Once present, the resistance trait can spread rapidly through bacterial populations under antibiotic selection pressure.
Several broad biological strategies allow bacteria to survive antibiotic exposure. A common mechanism involves modification of the drug’s molecular target. If a mutation alters the structure of the protein or enzyme that an antibiotic normally binds, the drug may no longer interact effectively with its target, reducing or eliminating its antimicrobial activity. Target modification underlies resistance to many drug classes, including macrolides, fluoroquinolones, and rifamycins.2
Another strategy reduces the amount of antibiotic that reaches its intended intracellular target. Some bacteria accomplish this by decreasing membrane permeability, limiting the entry of antibiotic molecules into the cell. Others rely on efflux pumps, membrane proteins that actively export antimicrobial compounds out of the cell before they can accumulate to toxic concentrations. Efflux-mediated resistance is particularly important in gram-negative bacteria, where multiple transport systems can contribute to multidrug resistance phenotypes.2
Bacteria may also neutralize antibiotics directly through enzymatic inactivation. Certain enzymes chemically modify or degrade antimicrobial molecules, rendering them ineffective. Beta-lactamases, which hydrolyze beta-lactam antibiotics, are among the best-known examples of this mechanism and have played a central role in the global spread of resistance to penicillins and related drugs.2
A final mechanism involves metabolic bypass pathways. In these cases, bacteria circumvent the antibiotic’s target biochemical step by activating alternative metabolic routes or acquiring replacement enzymes that perform the same function but are not inhibited by the drug. This strategy allows the organism to maintain essential cellular processes despite the presence of antimicrobial compounds.2
Each resistance mechanism alters core cellular processes in different ways, and the evolutionary costs of resistance vary widely depending on how the bacterium evades the antibiotic. The genetic changes required for resistance can alter cellular processes such as metabolism and membrane transport. As a result, different resistance strategies impose different physiological burdens on bacteria, which helps explain why some resistance traits persist even when antibiotics are absent, while others decline when selective pressure is removed.
Fitness Costs: Why Resistance Can Be Evolutionarily Expensive
The genetic mechanisms that allow bacteria to resist antibiotics frequently alter fundamental cellular processes. Because those processes are optimized by natural selection for efficient growth and survival, resistance mutations can disrupt normal physiology, thereby reducing bacterial fitness in the absence of antibiotics.
A common example involves antibiotics that target the bacterial ribosome. Mutations that modify ribosomal proteins or ribosomal RNA can prevent antibiotics from binding effectively, but those same alterations may also reduce the efficiency of protein synthesis. Slower or less accurate translation can limit bacterial growth rates, placing resistant strains at a competitive disadvantage in the absence of antibiotics.
Resistance mechanisms based on efflux pumps can impose different physiological burdens. These membrane proteins actively export antibiotics from the cell, but they require energy to function and can alter membrane transport dynamics. Increased expression of efflux pumps may divert limited metabolic resources from growth or disrupt the balance of molecules moving across the cell membrane, thereby putting cells at a growth disadvantage.
Changes in membrane permeability can produce additional tradeoffs. Some bacteria evolve resistance by modifying outer membrane proteins that normally allow antibiotics to enter the cell. While reduced permeability can limit antibiotic uptake, it may also restrict nutrient influx, thereby affecting metabolic efficiency and slowing bacterial replication.
These physiological effects illustrate why resistance mutations sometimes carry measurable fitness costs in antibiotic-free environments. When resistant strains grow more slowly or compete less effectively than susceptible strains, the evolutionary advantage conferred by resistance may diminish once antibiotic pressure declines.
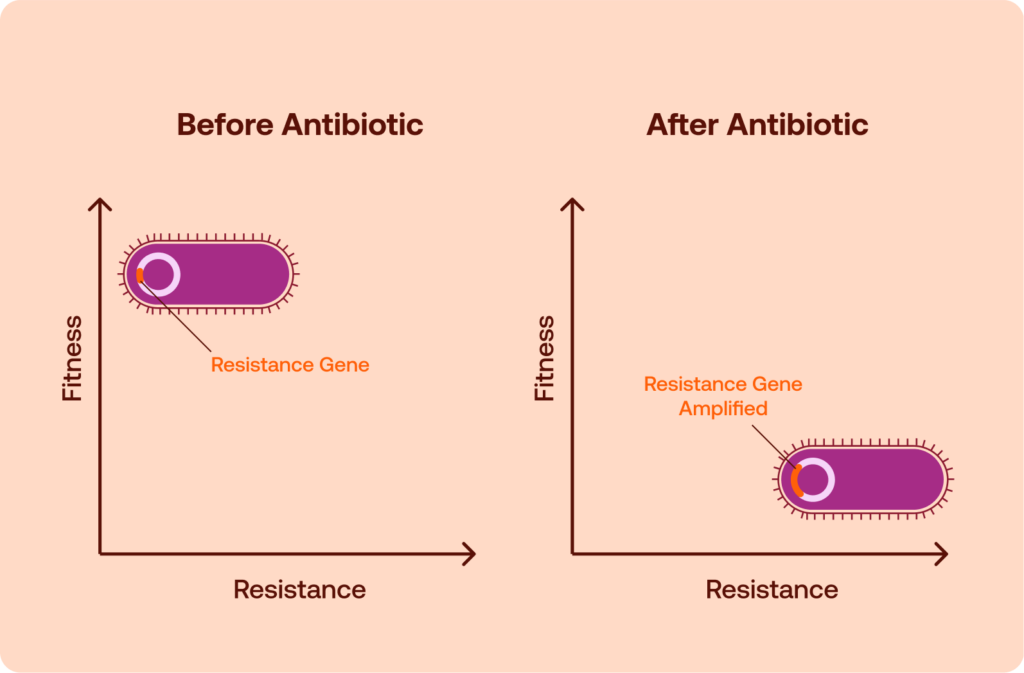
Why Resistance Does Not Always Disappear
Although resistance mutations can impose fitness costs, those costs do not necessarily persist over time. Bacterial populations evolve rapidly, and additional genetic changes can alter the evolutionary balance between resistant and susceptible strains. As a result, resistance traits that initially appear costly may remain stable or even become widespread long after antibiotic pressure has declined.
One important mechanism involves compensatory mutations. After an initial resistance mutation reduces bacterial fitness, subsequent mutations may restore normal physiological performance while preserving resistance. These compensatory changes can occur in the same gene that originally mutated or elsewhere in the genome, adjusting cellular pathways to offset the biological burden of resistance. Once such compensation occurs, resistant strains may regain competitive parity with susceptible bacteria, eliminating the evolutionary disadvantage that might otherwise have caused resistance to decline.3
The spread of resistance is also shaped by HGT, which allows bacteria to acquire genetic material from other organisms rather than inheriting it solely through vertical reproduction. Resistance genes can move between bacterial species through a range of mechanisms, such as conjugation, transformation, and transduction. Mobile genetic elements, including plasmids, transposons, and integrons, often carry multiple resistance genes and can disseminate them across microbial populations with remarkable efficiency.4
Plasmid-mediated resistance introduces additional evolutionary complexity. Many resistance determinants reside on plasmids, which replicate independently of the bacterial chromosome. Although plasmids can impose fitness costs on their host cells, they may persist through mechanisms such as high transfer rates between bacteria or compensatory adaptation that reduces their metabolic burden. This phenomenon has been described as the plasmid paradox: despite their potential cost, plasmids carrying resistance genes can remain stable within bacterial populations over long periods in the abscence of antibioticswit.5,6
Another factor that promotes persistence is co-selection. Resistance genes frequently cluster on the same mobile genetic elements, meaning that exposure to one antibiotic can simultaneously select for resistance to several others. In such cases, reducing the use of a single drug may have little effect if other antibiotics continue to select for the same genetic elements. Even when the original selective pressure disappears, continued exposure to unrelated antimicrobials can maintain resistance in the population.4
These evolutionary processes help explain why resistance does not always decline after antibiotic use decreases. Even when a resistance mutation initially imposes a measurable fitness cost, the evolutionary landscape surrounding that mutation can shift over time. Compensation, gene transfer, plasmid persistence, and co-selection all provide pathways through which resistance traits can stabilize within bacterial populations.
Real-world experience reflects this complexity. Some interventions that reduce antibiotic use have been followed by measurable declines in resistance, while others have produced little change. Understanding why those outcomes differ provides important insight into the evolutionary dynamics of antimicrobial resistance.
Natural Experiments in Resistance Reversal
If resistance carries evolutionary costs, reducing antibiotic use might allow susceptible strains to regain a competitive advantage. Several public health interventions have created real-world conditions in which this possibility could be evaluated. These natural experiments provide some of the clearest evidence about whether resistance can decline once selective pressure is reduced.
A widely cited example comes from Finland in the 1990s. In response to rising macrolide resistance among group A streptococci, national health authorities implemented a campaign to reduce macrolide use in outpatient settings. As prescribing declined, surveillance data revealed a corresponding reduction in erythromycin resistance among Streptococcus pyogenes isolates. The decrease in resistance followed the drop in antibiotic consumption, suggesting that reduced selective pressure allowed susceptible strains to re-establish themselves within the population.7
This case demonstrates that resistance reversal can occur under the right evolutionary conditions. When resistance imposes a biological cost, and antibiotic pressure decreases, susceptible bacteria may outcompete resistant ones. Over time, this shift in competitive dynamics can alter the composition of circulating bacterial populations.
However, not all attempts to reduce resistance by decreasing antibiotic use have yielded similar outcomes. A study in Sweden examined the effects of a large reduction in trimethoprim prescribing on resistance patterns in urinary Escherichia coli. Despite a substantial decline in trimethoprim use, researchers observed little evidence that trimethoprim resistance decreased in the bacterial population. Resistant strains remained common even after the reduction in antibiotic exposure.8
Investigators suggested several explanations for the persistence of resistance in this case. One possibility was that the resistance determinants involved imposed little or no measurable fitness cost, allowing resistant bacteria to remain competitive even in the absence of antibiotic pressure. Another factor was co-selection: trimethoprim resistance genes were often linked to resistance determinants for other antibiotics on the same mobile genetic elements. Continued exposure to those other drugs may have maintained selection for the entire resistance cluster, preventing trimethoprim resistance from declining.
These contrasting examples illustrate the complexity of resistance dynamics in real populations. Reducing antibiotic use can sometimes shift evolutionary pressures enough to allow susceptible strains to regain dominance. In other cases, compensatory evolution, gene linkage, or minimal fitness costs allow resistance traits to persist despite major changes in prescribing patterns. Reversal is therefore possible, but it cannot be assumed.
Antibiotic Cycling: A Strategy That Has Borne Few Fruits
The idea that resistance might decline when selective pressure is reduced led to efforts to deliberately manipulate patterns of antibiotic use. One of the most widely discussed approaches has been antibiotic cycling, a stewardship strategy in which hospitals periodically rotate the primary antibiotic classes used for specific infections. The rationale is straightforward: by regularly changing the dominant therapy, clinicians might prevent bacteria from experiencing prolonged exposure to any single drug, reducing the likelihood that resistant strains will dominate.
In theory, cycling could disrupt the evolutionary advantage of resistant organisms. If a single antibiotic class is used heavily for an extended period, resistant strains may emerge or expand. Rotating to a different drug class might remove the selective advantage associated with the first antibiotic, potentially allowing susceptible strains to recover while selecting against organisms resistant to the newly introduced therapy. The concept attracted considerable attention because it seemed to offer a practical way to manage resistance through prescribing policies.
In practice, however, the results have been inconsistent. Studies examining antibiotic cycling in hospital settings have produced mixed findings, with some reporting modest improvements in resistance patterns and others finding little or no effect. Differences in study design, patient populations, local microbial ecology, and prescribing practices have made it difficult to draw clear conclusions from individual trials.
A systematic review of cycling interventions in hospitals illustrates the challenge. The analysis found that existing studies produced conflicting results and that the available evidence did not demonstrate that cycling strategies were superior to alternative approaches, such as antibiotic mixing, in which multiple drug classes are used concurrently across patients rather than rotated over time.9 The authors also noted the limited number of rigorous randomized studies evaluating these strategies.
These findings suggest that simple rotation schemes do not reliably alter the evolutionary dynamics of bacterial populations. Resistance evolution depends on a complex interaction of genetic mechanisms, microbial ecology, and antibiotic exposure patterns. Changing prescribing practices on a fixed schedule may not be sufficient to overcome those underlying forces.
Collateral Sensitivity: The Evolutionary Trade-Off Opportunity
The mixed results of antibiotic cycling reflect a broader reality: bacterial evolution does not respond predictably to simple prescribing patterns. However, research in evolutionary microbiology has identified a more structured phenomenon that may offer a way to guide resistance dynamics more deliberately. This concept, known as collateral sensitivity, arises when a genetic change that confers resistance to one antibiotic simultaneously increases susceptibility to another.
Experimental evolution studies have demonstrated that the fitness trade-offs inherent in bacterial adaptation can recur during the evolution of resistance. Laboratory experiments in which bacterial populations are exposed to gradually increasing antibiotic concentrations have shown that resistance mutations often generate predictable patterns of cross-resistance and collateral sensitivity across multiple drugs. These relationships can form networks in which adaptation to one antibiotic alters susceptibility profiles across several others.10
To map these relationships, researchers often use experimental evolution platforms in which bacterial populations are exposed to controlled sequences of antibiotics over many generations. By repeatedly selecting for resistance and measuring resulting susceptibility profiles, scientists can construct resistance landscapes that describe how adaptation to one drug alters sensitivity to others. These landscapes reveal that resistance evolution often follows pathways constrained by underlying physiological trade-offs. In some cases, the order in which antibiotics are applied can influence which evolutionary routes bacteria take, creating opportunities to steer populations toward states that remain vulnerable to certain drugs. This concept — sometimes described as evolutionary steering — suggests that treatment sequences may be designed not only to cure the immediate infection but also to shape the resistance trajectories that follow.
Such patterns have important implications for antibiotic stewardship. If resistance to one drug increases vulnerability to another, carefully designed treatment sequences could potentially exploit these evolutionary constraints. Instead of selecting antibiotics solely based on immediate clinical effectiveness, therapy could be structured to steer bacterial populations toward evolutionary states that remain susceptible to future treatments.
The concept shifts the focus from simply suppressing resistance to actively shaping the evolutionary trajectory of bacterial populations. In principle, a sequence of antibiotics chosen to exploit collateral sensitivity relationships could limit the long-term accumulation of resistance. By anticipating the trade-offs associated with adaptation, clinicians might be able to guide bacterial evolution toward less dangerous outcomes rather than allowing it to proceed unchecked.
Limits And Instability of Collateral Sensitivity
Although collateral sensitivity offers an appealing framework for steering bacterial evolution, its practical application is constrained by several biological complexities. The tradeoffs that emerge during resistance evolution are not uniform across organisms, drugs, or genetic pathways. As a result, collateral sensitivity patterns observed in one bacterial species or experimental system may not appear in another.
One source of variability lies in the diversity of resistance mechanisms themselves. Different mutations can produce distinct physiological consequences even when they confer resistance to the same antibiotic. Because collateral sensitivity arises from those underlying physiological changes, the specific mutation that evolves can determine whether a vulnerability to another drug appears at all. Experimental studies have shown that evolutionary trajectories during resistance development can diverge across replicate populations, producing different patterns of collateral sensitivity or cross-resistance depending on which mutations arise first.11,12
Collateral sensitivity relationships can also vary across bacterial species. Differences in cell structure, metabolic networks, and drug transport systems influence how resistance mutations affect cellular physiology. As a result, trade-offs observed in one organism may not translate directly to others. Comparative studies examining multiple bacterial species have found that collateral sensitivity and cross-resistance patterns differ substantially across organisms, reflecting the diversity of resistance mechanisms and cellular responses to antibiotic stress.13
Another challenge involves the evolutionary stability of these tradeoffs. Even when collateral sensitivity initially emerges, continued adaptation may erode the vulnerability. Bacterial populations can acquire additional mutations that compensate for the sensitivity while maintaining resistance to the original drug. Over time, this process can transform an initially favorable tradeoff into broader cross-resistance, undermining treatment strategies that rely on predictable evolutionary relationships.14
These findings highlight the importance of approaching collateral sensitivity with caution; it represents a promising conceptual tool for guiding antibiotic strategies, but it cannot yet be treated as a universally stable property of resistance evolution.
Toward Evolutionary Guidance Strategies
The limitations of both simple antibiotic withdrawal and fixed cycling schemes point toward a more nuanced approach to stewardship that treats antimicrobial resistance as an evolutionary system that can potentially be guided rather than merely reacted to. Instead of assuming that resistance will fade once antibiotic pressure is removed or attempting to suppress resistance through rigid rotation schedules, this perspective focuses on shaping the selective pressures that bacterial populations experience over time.
At the center of this approach is the idea of evolution-guided stewardship. The goal is not simply to reduce antibiotic use but to manage how different drugs influence the evolutionary trajectory of bacterial populations. In this framework, antibiotic selection and sequencing are informed by knowledge of resistance mechanisms, collateral sensitivity relationships, and the genetic pathways through which bacteria typically adapt to antimicrobial exposure.
In practice, such an approach could resemble a more adaptive form of antimicrobial stewardship. Treatment decisions might incorporate pathogen-specific resistance mechanisms identified through genomic diagnostics, allowing clinicians to select antibiotics that exploit known evolutionary tradeoffs. Regional surveillance data could help identify which resistance pathways dominate in which settings, enabling stewardship programs to adjust recommended therapies accordingly. Over time, computational models or machine learning tools might assist clinicians in choosing drug sequences that minimize the probability of resistance emergence while preserving the effectiveness of existing antibiotics.
Improved surveillance of resistance mechanisms may be critical to enable such strategies. Modern genomic tools make it increasingly possible to track not only which pathogens are resistant but also which specific genetic determinants underlie that resistance. Monitoring those mechanisms can provide insight into the evolutionary pressures shaping local microbial populations and help identify opportunities where resistance tradeoffs might be exploited.
Another element involves a strategic sequencing of antibiotics. Rather than choosing treatments independently for each infection, clinicians could consider how one antibiotic might influence the subsequent susceptibility profile. In systems where collateral sensitivity relationships are known, a sequence of drugs could be designed to steer bacterial populations toward evolutionary states that remain vulnerable to future therapies.
Experimental studies have shown that resistance evolution can create predictable patterns of susceptibility across multiple antibiotics. Mapping these collateral-sensitivity relationships raises the possibility of designing treatment sequences that deliberately exploit evolutionary trade-offs, thereby limiting the long-term accumulation of resistance.
Targeted reductions in the use of specific drug classes could also play a role. When resistance mechanisms impose meaningful fitness costs, decreasing selective pressure on a particular antibiotic may create conditions in which susceptible strains regain a competitive advantage. Combined with strategic sequencing and surveillance, such reductions could form part of a broader effort to reshape microbial populations rather than simply responding to resistance after it emerges.
Designing these strategies requires careful attention to the dynamics of evolution itself. The effectiveness of antibiotic sequences depends on factors such as the order in which drugs are used, the biology of the pathogen involved, and the evolutionary pathways available to resistant organisms.
This evolutionary perspective does not offer a simple solution to antimicrobial resistance. Instead, it suggests that stewardship strategies must account for the adaptive behavior of microbial populations. By integrating surveillance, evolutionary biology, and clinical practice, it may be possible to move beyond reactive approaches and begin shaping the selective environments that drive resistance in the first place.
Policy Implications and Research Priorities
Moving from theoretical models of evolutionary stewardship to practical implementation would require substantial advances in surveillance, data integration, and clinical research. The central challenge is understanding how resistance emerges, spreads, and evolves over time within specific microbial populations. Without reliable information about those dynamics, attempts to guide the evolution of resistance would amount to little more than educated guesswork.
High-resolution resistance surveillance is therefore a foundational requirement. Traditional antimicrobial susceptibility testing identifies whether pathogens are resistant to specific drugs, but evolutionary strategies demand more granular insight into how resistance is developing within a population. Monitoring changes in resistance prevalence over time, across geographic regions, and within specific healthcare settings would provide the baseline data needed to detect evolutionary shifts.
Genomic monitoring of resistance mechanisms represents a critical extension of that surveillance. Advances in microbial genomics have made it possible to identify the precise genetic determinants responsible for antibiotic resistance. Tracking those determinants allows researchers and clinicians to distinguish between different resistance pathways that may have distinct evolutionary consequences. Because collateral sensitivity and fitness costs depend on the underlying mechanisms of resistance, genomic data can help identify which bacterial populations may be most responsive to evolutionary stewardship strategies.
Clinical trials would also be necessary to determine whether evolutionary guidance can produce measurable benefits in real-world settings. Laboratory evolution studies have demonstrated that antibiotic sequences can influence resistance trajectories under controlled conditions. Translating those findings into clinical practice would require trials to evaluate whether treatment regimens designed around evolutionary principles can reduce the emergence of resistance, preserve antibiotic effectiveness, or improve patient outcomes.
Any such strategies would need to operate within existing antimicrobial stewardship frameworks. International initiatives, such as the World Health Organization’s Access, Watch, and Reserve (AWaRe) classification, already emphasize monitoring antibiotic use, promoting appropriate prescribing practices, and evaluating the impact of stewardship policies on resistance patterns. These programs focus on optimizing antibiotic use and improving surveillance rather than relying on blanket withdrawal policies, providing a policy framework in which more sophisticated evolutionary approaches could be tested and implemented.15
Evolution-guided stewardship would require coordination across microbiology laboratories, clinical researchers, public health agencies, and healthcare systems. The same infrastructure that supports modern antimicrobial stewardship programs could also provide the foundation for evaluating whether evolutionary strategies can help preserve antibiotic effectiveness over the long term.
Approaching stewardship through an evolutionary lens does not guarantee that resistance can be reversed or controlled. It does suggest that the selective environments created by antibiotic use can influence how resistance emerges and spreads. Designing stewardship strategies that deliberately account for those dynamics may offer one of the most promising avenues for preserving the effectiveness of antibiotics in the decades ahead.
In August 2016, physicians at a hospital in Reno, Nevada, faced a scenario that infectious disease specialists had long warned about but hoped never to see. A woman in her seventies had been admitted with a severe bacterial infection after receiving medical treatment overseas. Doctors attempted antibiotic after antibiotic, moving through the entire arsenal available in the United States.
None worked.
Laboratory analysis revealed that the Klebsiella pneumoniae strain infecting her carried resistance mechanisms that rendered every available antimicrobial drug ineffective. The infection proved untreatable, and the patient ultimately died of septic shock.¹ Further testing confirmed that the organism was resistant to all 26 antibiotics examined, including drugs typically reserved as last-line treatments.²
Cases like this were once considered extreme anomalies. Now, antimicrobial resistance (AMR) is widely recognized as one of the most significant global public health challenges of the twenty-first century.³ A comprehensive global analysis estimated that drug-resistant bacterial infections were responsible for 1.27 million deaths worldwide in 2019, while 4.95 million deaths were associated with resistant infections.⁴ These numbers place AMR among the leading causes of death globally and underscore a rapidly growing threat to modern medicine.
Antibiotics transformed healthcare in the twentieth century, turning infections that once killed millions into treatable conditions. That foundation, however, is beginning to weaken.
The Biology of Resistance
Antibiotics work by targeting essential biological processes inside bacterial cells. Some disrupt the synthesis of bacterial cell walls, causing cells to rupture. Others interfere with protein production or DNA replication, preventing bacteria from multiplying.
But bacterial populations evolve quickly. When exposed to antibiotics, most bacteria die, yet some may possess genetic traits that allow them to survive. Those survivors reproduce, passing their resistance traits to future generations.
Over time, this evolutionary process allows resistant strains to dominate microbial populations exposed to antibiotics.
Bacteria have evolved numerous strategies to evade antimicrobial drugs. Some produce enzymes that break down antibiotics before they reach their targets. Others modify the molecular structures that antibiotics are designed to attack. Still others actively pump antimicrobial compounds out of the cell before they reach lethal concentrations.
And as if family-line mutations weren’t enough of a challenge, resistance traits can also spread rapidly within microbial communities. Bacteria can acquire resistance genes through mutation or by mechanisms that enable them to obtain DNA from neighboring microbes. This capacity rapidly accelerates the spread of resistance traits across bacterial populations and environments, especially in settings where multiple ill patients are in close proximity (e.g., hospitals, nursing homes).

The result of a microbe’s proclivity for mutation is a continuous evolutionary arms race between microbes and medicine. Each new antibiotic exerts selective pressure that favors organisms capable of surviving it. As these organisms proliferate, treatments that once worked reliably become progressively less effective.
The Quiet Erosion of Modern Medicine
The consequences of antimicrobial resistance extend far beyond microbiology laboratories. Many of the medical procedures that define modern healthcare depend on antibiotics functioning reliably.
Routine surgeries rely on prophylactic antibiotics to prevent infection during or after procedures. Chemotherapy suppresses immune function, leaving patients dependent on antimicrobial drugs to prevent life-threatening infections. Organ transplantation requires long-term immunosuppression, which similarly increases vulnerability to bacterial disease.
When antibiotics fail, these procedures become significantly more dangerous.
Hospital-acquired infections caused by resistant organisms already pose serious challenges in healthcare systems worldwide. Pathogens such as Escherichia coli, Klebsiella pneumoniae, Staphylococcus aureus, and Acinetobacter baumannii account for a substantial share of global deaths linked to antimicrobial resistance.⁵
Infections caused by these organisms often require prolonged hospitalization and complex treatment regimens. When first-line antibiotics fail, clinicians may be forced to rely on older drugs that are less effective or more toxic.
This quiet erosion of antibiotic effectiveness rarely makes headlines, yet it is gradually reshaping everyday medical practice.
A Post-Antibiotic Era
Public health experts frequently warn of the possibility of a “post-antibiotic era,” a future in which common infections once again become difficult or impossible to treat.
The phrase may sound dramatic, but it reflects a trajectory already visible in isolated cases.
The Nevada infection illustrates the point starkly. The bacterium responsible for the patient’s illness carried resistance mechanisms that neutralized every antimicrobial drug available to physicians.¹
Although infections that resist all available antibiotics remain uncommon, they represent the logical endpoint of the evolutionary processes driving antimicrobial resistance.
If such pathogens become widespread, the implications for modern medicine could be profound.
Surgical procedures could become far more dangerous due to increased infection risk. Cancer treatments that suppress immune function could expose patients to infections that medicine can no longer control. Organ transplantation programs could face severe limitations because immunosuppressed patients would be highly vulnerable to untreatable infections.
Even infections that were once considered routine, including pneumonia or urinary tract infections, could again become major causes of death.
Modeling studies suggest that antimicrobial resistance could cause tens of millions of deaths globally in the coming decades if current trends continue without significant intervention.⁶
These projections underscore the urgency of addressing AMR before such scenarios become commonplace.
Why Resistance Is Accelerating
Several forces have contributed to the rapid emergence and spread of antimicrobial resistance over the past several decades.
One major driver is the widespread use of antibiotics in human medicine. While these drugs are essential tools for treating bacterial infections, they are sometimes prescribed unnecessarily, such as for viral illnesses where they offer no benefit.
Incomplete treatment courses can also contribute to resistance. When patients stop taking antibiotics before the full course is complete, partially resistant bacteria may survive and multiply.
Healthcare environments themselves can facilitate the spread of resistant organisms. Hospitals concentrate vulnerable patients, invasive procedures, and antimicrobial drugs in a single setting, creating conditions that allow resistant pathogens to emerge and spread.
Environmental factors may also play a role. Antibiotics released into wastewater from healthcare facilities, agriculture, or pharmaceutical manufacturing can expose environmental bacteria to low levels of antimicrobial compounds, encouraging the development of resistance.
Together, these pressures create a global microbial ecosystem in which resistant bacteria can thrive.
The Global Response
In recognition of the dire concern that scientists, public health officials, and other leaders have about the effects of antimicrobial resistance on our future, significant efforts are underway to combat AMR.
International health organizations and national public health agencies have established surveillance systems to monitor resistance patterns worldwide. These systems collect data on resistant infections, helping researchers identify emerging threats and guide policy responses.
Hospitals and healthcare systems are also increasingly implementing antimicrobial stewardship programs. These initiatives aim to ensure that antibiotics are used only when necessary and that the most appropriate drugs are selected for each infection.
Researchers and biotechnology companies are simultaneously working to develop new antimicrobial therapies. However, antibiotic development faces unique economic challenges. Unlike many other drugs, antibiotics are typically used for short treatment courses and are often reserved for severe infections, limiting their commercial revenue generation opportunities.
To address this issue, new funding models and public-private partnerships have emerged to support antimicrobial research and development.
Scientists are also exploring alternative approaches to combating bacterial infections, including bacteriophage therapies, antimicrobial peptides, and treatments that neutralize bacterial virulence factors rather than directly killing the organisms.
Advances in diagnostic technology are also helping clinicians identify infections more quickly and determine which antibiotics are most likely to be effective, reducing unnecessary antibiotic use and enabling more targeted treatments.
An Evolutionary Arms Race
Despite the role of modern medicine and other global industries in exacerbating the AMR crisis, antimicrobial resistance ultimately stems from evolutionary biology. Microorganisms evolve rapidly, and the selective pressures created by widespread antibiotic use ensure that resistance will continue to emerge.
The challenge facing global health systems is therefore not to eliminate resistance entirely, but to manage it. Doing so will require a combination of scientific innovation, responsible antibiotic use, improved infection prevention, and coordinated global action.
Vaccination programs, sanitation improvements, and infection control measures can all reduce the number of infections that require antibiotic treatment in the first place. Meanwhile, new antimicrobial therapies and diagnostics will be needed to keep pace with evolving pathogens.
The microbes humanity is fighting have had billions of years to refine their survival strategies. Antibiotics have been part of human medicine for barely a century.
Whether the next century belongs to resistant pathogens or to a new generation of antimicrobial innovation will depend on how effectively science, healthcare systems, and global policy adapt to this rapidly evolving challenge. disAMR is dedicated to fostering exposure, funding, collaboration, and persistence necessary to counter antimicrobial resistance. Will you join us? Add your name to the list to receive updates and information.
Introduction — The Hidden Vulnerabilities of Bacteria
Antimicrobial drugs succeed because bacteria, despite their remarkable adaptability, rely on biological systems that differ in important ways from those of human cells. These differences create opportunities for selective intervention. An antimicrobial compound that interferes with a process unique to bacteria can cripple a pathogen while leaving human tissues largely unaffected. This fundamental principle underlies the field of antibacterial therapy and explains why antibiotics have remained such powerful tools in medicine for more than 80 years.
A single microbial cell operates an intricate network of molecular machinery that maintains its structure, produces proteins, replicates its genetic material, and generates the metabolic intermediates needed for growth and division. Most antibiotics work by interfering with an activity essential to the survival of the pathogenic bacteria.
Viewed conceptually, antimicrobial therapy resembles sabotage inside a factory. A drug might disable the workers building the outer wall, jam the assembly line that produces proteins, damage the copying machines that reproduce genetic instructions, or block the supply chains that deliver essential biochemical components. When any of these core systems fails, bacterial growth slows or stops, and in many cases, the cell dies.
Although hundreds of antibiotic molecules have been discovered, most act through a limited number of underlying strategies. These mechanisms fall into several broad categories: inhibition of cell wall construction, disruption of protein synthesis, interference with DNA or RNA production, blockade of key metabolic pathways, and damage to the bacterial membrane itself. Understanding these mechanisms provides a useful framework for exploring both the strengths of existing antimicrobial drugs and the challenges posed by the evolution of resistance.

Breaking the Walls: Cell Wall Synthesis Inhibitors
For many bacteria, survival depends on the integrity of the cell wall, a rigid mesh-like shell surrounding the membrane that protects the cell from osmotic pressure. Because the concentration of dissolved molecules inside the cell is often higher than in the surrounding environment, water tends to flow inward. The cell wall provides the mechanical strength needed to prevent swelling and rupture, allowing bacteria to maintain their shape and structural stability.
The wall itself is built primarily from peptidoglycan, a lattice of sugar chains connected by short peptide bridges. These cross-linked strands create a resilient network analogous to reinforced concrete. Because human cells lack this structure entirely, the enzymes responsible for assembling peptidoglycan are attractive, highly selective targets for antimicrobial drugs.
Among the most important cell wall inhibitors are the β-lactam antibiotics, a family that includes penicillins, cephalosporins, carbapenems, and related compounds. These drugs act by interfering with the final stages of peptidoglycan assembly. During normal cell wall construction, penicillin-binding proteins (PBPs) connect peptide strands to create the cross-linked framework that gives the wall its strength. β-lactams bind covalently to PBPs, inhibiting their transpeptidase activity and preventing cross-linking of peptidoglycan strands. As a result, newly synthesized cell wall material becomes structurally weak and unstable.1
The consequences for the bacterium are often catastrophic. As the cell continues to grow and divide, defects accumulate in the wall. Eventually, the weakened structure can no longer withstand internal pressure, leading to rupture of the cell membrane and the organism’s death. For this reason, β-lactam antibiotics are typically bactericidal rather than merely bacteriostatic. Their ability to kill bacteria directly and relatively low toxicity to human cells have made them one of the most widely used and clinically successful antibiotic classes in medical history.1
Despite their effectiveness, β-lactams are also vulnerable to resistance. Many bacteria produce β-lactamases, enzymes that chemically degrade the antibiotic before it can reach its target. Others acquire altered PBPs that bind β-lactams less effectively while still carrying out cell wall synthesis. These adaptations allow pathogens to preserve the essential cross-linking process even in the presence of the drug.2
A second group of cell wall inhibitors takes a different approach to the same biological system. Glycopeptide antibiotics, such as vancomycin, do not disable the enzymes responsible for assembling the wall. Instead, they bind directly to the molecular building blocks used to construct it. During peptidoglycan synthesis, precursor molecules containing a terminal d-alanine–d-alanine (d-Ala–d-Ala) motif are incorporated into the growing structure. Vancomycin binds tightly to this motif, preventing precursors from being incorporated into the cell wall lattice.3
This strategy effectively removes the raw materials needed for construction. While the enzymes responsible for assembly remain functional, they cannot access the substrates required to extend the wall. As a result, bacterial growth stalls and the structural integrity of the cell envelope gradually fails. Glycopeptides have therefore become particularly important for treating infections caused by Gram-positive organisms, including pathogens that are resistant to many other antibiotics.
Resistance to glycopeptides arises through a different mechanism than resistance to β-lactams. Instead of destroying the drug or altering an enzyme target, some bacteria modify the structure of the cell wall precursor itself. In certain resistant strains, the d-Ala–d-Ala motif is replaced with d-Ala–d-Lac. This subtle chemical change reduces vancomycin binding dramatically while still allowing peptidoglycan assembly to proceed.4
Sabotaging the Protein Factory: Ribosome Inhibitors
If the cell wall provides structural protection, proteins provide function. Enzymes catalyze metabolic reactions, structural proteins maintain cellular organization, and regulatory proteins coordinate responses to environmental changes. Nearly every activity required for bacterial survival depends on the continuous production of new proteins. Interrupting this process quickly slows growth and, in some cases, kills the cell outright.
Protein production takes place in ribosomes, molecular complexes that translate genetic instructions encoded in messenger RNA (mRNA) into chains of amino acids. Bacterial ribosomes differ structurally from their human counterparts, allowing drugs to interfere with microbial protein synthesis without disrupting the same process in human cells. For this reason, the ribosome has been one of the most productive targets in antimicrobial therapy.
Several antibiotic families interfere with protein synthesis, each exploiting a different vulnerability in the ribosome’s operation. Some distort the translation process itself, while others physically block the movement of molecular components needed for protein assembly.
Aminoglycoside drugs bind to the 30S ribosomal subunit and interfere with the decoding of mRNA. Under normal circumstances, the ribosome reads genetic instructions three nucleotides at a time, matching each codon to the corresponding transfer RNA (tRNA) that carries a specific amino acid. Aminoglycosides disrupt this proofreading process, causing the ribosome to misread the message. The resulting proteins contain incorrect amino acids and often fail to fold properly or perform their intended functions.5,6
The accumulation of defective proteins can damage cellular processes in several ways. Some proteins lose their enzymatic activity, while others become unstable or interfere with essential pathways. In severe cases, the errors introduced during translation lead to the production of toxic proteins that compromise membrane integrity or other critical systems. Because of these cascading effects, aminoglycosides often display rapid bactericidal activity rather than merely slowing bacterial growth.6
Bacteria can resist aminoglycosides through several mechanisms, but one of the most common involves enzymatic modification of the drug itself. Certain bacterial enzymes chemically alter the antibiotic molecule through processes such as acetylation, phosphorylation, or adenylation. These modifications reduce the drug’s ability to bind the ribosome, allowing protein synthesis to continue even in the presence of the antibiotic.
Macrolide antibiotics take a different approach to disrupting protein production. Instead of causing decoding errors, these drugs physically obstruct part of the ribosome’s architecture. Macrolides bind to the large, 50S ribosomal subunit within a channel known as the nascent peptide exit tunnel. As new proteins are synthesized, the growing amino acid chain normally passes through this tunnel and emerges from the ribosome. When a macrolide occupies this space, the passage becomes partially blocked, preventing the newly forming peptide from extending properly.7
This interference stalls the ribosome during translation, halting protein synthesis before the molecule is complete. Because macrolides act at a defined structural site within the ribosome, relatively small changes to that site can confer resistance. A common mechanism involves methylation of the ribosomal RNA that lines the exit tunnel. This chemical modification alters the shape of the binding pocket, reducing macrolides’ ability to bind to the ribosome.8
A third group of ribosome-targeting antibiotics, the tetracyclines, disrupts protein synthesis at an earlier step in the process. These drugs bind to the 30S subunit at a region known as the A-site, where incoming tRNA molecules normally deliver amino acids to the ribosome. By occupying this position, tetracyclines prevent aminoacyl-tRNA from entering the ribosome and participating in translation.9
Without a steady supply of tRNA molecules bearing amino acids, translation stalls, and the bacterium is unable to produce the proteins required for metabolism, replication, and structural maintenance. Tetracyclines therefore act primarily as bacteriostatic agents, slowing bacterial growth and allowing the immune system to clear the infection.
Resistance to tetracyclines often arises through one of two strategies. Efflux pumps embedded in the bacterial membrane can actively export the drug from the cell, reducing its intracellular concentration. Alternatively, specialized ribosomal protection proteins can dislodge tetracyclines from the ribosome, restoring the ability of tRNA molecules to bind and resume translation.10
Disrupting the Blueprint: DNA and RNA Inhibitors
Bacteria depend on accurate transmission of genetic information, which requires DNA replication to copy the genome and transcription to produce messenger RNA (mRNA) that guides protein synthesis. Several antibiotic classes exploit this vulnerability by interfering with the enzymes responsible for these processes.
Fluoroquinolones target the machinery that allows bacterial chromosomes to be replicated and organized. As a circular DNA molecule is copied, it becomes tightly twisted and tangled. Specialized enzymes known as DNA gyrase and topoisomerase IV relieve this mechanical strain by cutting the DNA, allowing it to unwind, and then resealing the strands. Fluoroquinolones bind to these enzymes and stabilize an intermediate stage in this process in which the DNA has been cleaved but not yet rejoined.11
This stabilization traps the enzyme–DNA complex in a dysfunctional state. Instead of relieving tension in the chromosome, the enzyme remains locked onto broken DNA strands, preventing replication from proceeding. As these stalled complexes accumulate, double-strand DNA breaks can form, leading to lethal damage within the bacterial genome.12 Because these enzymes play such central roles in chromosome replication and organization, inhibiting them can halt bacterial growth across a wide range of species. Fluoroquinolones thus exhibit broad-spectrum antibacterial activity and have been widely used to treat infections of the respiratory and urinary tracts and other systems.
Resistance to fluoroquinolones most often arises through mutations in the genes encoding DNA gyrase or topoisomerase IV. These mutations subtly alter the enzyme’s structure, causing the antibiotic to bind less effectively while the enzyme continues to perform its normal function. In addition, some bacteria employ efflux pumps that actively export fluoroquinolones from the cell, reducing the concentration of the drug available to interact with the target enzymes.
While fluoroquinolones disrupt DNA replication, rifamycins interfere with transcription by binding to the β-subunit of bacterial RNA polymerase. Once attached, the drug blocks the extension of newly formed RNA chains, effectively preventing the transcription process from progressing beyond its earliest steps.13
Without the ability to produce RNA transcripts, bacteria cannot generate the mRNA molecules required for protein synthesis. The entire chain of information flow — from DNA to RNA to protein — breaks down. Rifamycins therefore exert a powerful antibacterial effect and have become central components of treatment regimens for diseases such as tuberculosis.
Resistance to rifamycins typically arises from mutations in the rpoB gene, which encodes the RNA polymerase β-subunit. These mutations alter the antibiotic’s binding site, reducing its ability to interact with the enzyme while leaving the transcription machinery functional.14 This mechanism illustrates a recurring theme in antimicrobial resistance: small genetic changes in a drug target can allow bacteria to evade inhibition while preserving essential cellular processes.
Starving the Cell: Metabolic Pathway Inhibitors
Not all antibiotics act by damaging major cellular structures or halting the production of proteins and nucleic acids. Some instead target the metabolic pathways that supply bacteria with the molecules required to sustain growth. These pathways function as biochemical supply chains, producing the building blocks needed for DNA synthesis, amino acid metabolism, and other essential cellular processes. Interrupting one of these pathways deprives the bacterium of key compounds, slowing or stopping its ability to replicate.
One of the best-known examples involves the folate biosynthesis pathway. Folate molecules play a central role in the synthesis of nucleotides. Without a reliable supply of folate, bacteria cannot produce the genetic material needed for cell division. Unlike humans, who obtain folate from dietary sources, many bacteria must synthesize it internally, providing a selective opportunity for antimicrobial therapy.
Two closely related drug classes exploit this pathway: sulfonamides and trimethoprim. Sulfonamides interfere with an early step in folate production by inhibiting dihydropteroate synthase, which normally helps assemble the precursor molecules that ultimately lead to the formation of dihydrofolate, an intermediate in the folate pathway. When sulfonamides block this step, the bacterium cannot produce sufficient folate to support DNA synthesis.15
Trimethoprim acts further downstream in the same pathway. It inhibits dihydrofolate reductase, the enzyme that converts dihydrofolate to tetrahydrofolate, the active form required for nucleotide synthesis and other metabolic reactions.16 When this conversion is blocked, the folate cycle stalls, and the cell is unable to produce the nucleotides needed for replication.
Because these drugs target sequential steps in the same pathway, they are often used together in combination therapy. The paired inhibition creates a metabolic bottleneck that is far more difficult for bacteria to bypass than either drug alone would be. This synergy enables the combination to achieve greater antibacterial activity while reducing the likelihood of resistance arising from simple alterations at a single enzyme target.
Attacking the Membrane: Last-Line Agents
Beyond the cell wall lies another critical defensive structure: the bacterial membrane. This lipid-based barrier encloses the cytoplasm and regulates the movement of molecules into and out of the cell. In gram-negative bacteria, the membrane system is even more complex, consisting of both an inner membrane and an outer membrane that contains lipopolysaccharide (LPS). Together, these layers create a formidable protective envelope that shields the bacterium from environmental stress and many antimicrobial agents. Because membrane integrity is essential for survival, drugs that damage this structure can rapidly compromise bacterial viability.
Polymyxins exploit this vulnerability. These molecules are cyclic peptides with a strong affinity for lipid A, a component of LPS that lines the outer membrane of Gram-negative bacteria. Under normal conditions, lipid A molecules are stabilized by interactions with divalent cations that help maintain the tight packing of the membrane surface. Polymyxins displace these stabilizing ions and bind directly to lipid A, disrupting membrane organization.17
This disruption weakens the outer membrane and increases its permeability. As the membrane structure deteriorates, essential cellular contents can leak out, and toxic molecules can enter. The resulting damage interferes with multiple cellular processes and ultimately leads to bacterial death. Because the mechanism involves direct destabilization of the membrane rather than inhibition of a specific enzyme, polymyxins can remain effective against gram-negative pathogens that resist many other antibiotics.
For this reason, polymyxins are often reserved as last-line treatments for severe infections caused by multidrug-resistant organisms, including difficult pathogens, such as Acinetobacter baumannii, Pseudomonas aeruginosa, and certain strains of Klebsiella pneumoniae. Their ability to breach the outer membrane makes them particularly valuable in situations where other drugs cannot reach their intracellular targets.
Resistance to polymyxins typically arises through modifications of the bacterial outer membrane itself. Some bacteria alter the chemical structure of lipid A, reducing polymyxins’ ability to bind effectively. These modifications alter the membrane surface charge, weakening the electrostatic interactions that normally attract the antibiotic molecule.18 Although such resistance mechanisms are still relatively uncommon compared with resistance to older antibiotic classes, their emergence highlights the continuing evolutionary pressure faced by even the most potent antimicrobial agents.
Why Resistance Is Inevitable
The remarkable adaptability of bacteria constantly challenges the effectiveness of antimicrobial drugs. Microbial populations reproduce quickly, and each generation introduces opportunities for genetic variation. Within these vast populations, even rare mutations can provide a survival advantage when antibiotics are present. Over time, natural selection favors organisms that can withstand the drug’s effects, allowing resistant strains to expand while susceptible ones disappear.
Mutation is not the only path through which resistance emerges. Bacteria can also acquire resistance genes from other organisms through a process known as horizontal gene transfer. Unlike vertical inheritance, in which genetic changes pass from parent cells to their descendants, horizontal gene transfer allows bacteria to exchange genetic material directly across species and even across genera. Resistance genes are frequently carried on plasmids — small, circular DNA molecules that replicate independently of the bacterial chromosome — or on mobile genetic elements such as transposons that can move between different DNA molecules. When these elements carry genes encoding β-lactamases, efflux pumps, or modified drug targets, they can spread resistance traits rapidly through microbial populations. In environments where antibiotics are present, this genetic exchange accelerates the emergence of multidrug-resistant strains and enables resistance mechanisms discovered in one species to disseminate across many others rapidly.
Several well-established mechanisms enable bacteria to evade antimicrobial activity. One common strategy involves producing enzymes that chemically destroy or modify the antibiotic before it reaches its target. β-lactamases provide a classic example: these enzymes break the β-lactam ring structure of penicillins and related drugs, rendering them inactive. Similar enzymatic processes can alter other antibiotic classes, preventing the drug from binding effectively to its intended target.19
Another route to resistance involves changes to the molecular structures that antibiotics normally attack. If a mutation alters the shape of a target protein or enzyme, the drug may no longer bind tightly enough to inhibit its function. The bacterium retains the ability to perform the essential cellular process while avoiding the drug’s inhibitory effects. Mutations in penicillin-binding proteins, ribosomal RNA, or DNA gyrase illustrate how small genetic changes can produce large shifts in antibiotic susceptibility.
Bacteria can also prevent antibiotics from reaching their targets in the first place. Efflux pumps embedded in the cell membrane actively transport certain drugs out of the cell, lowering intracellular concentrations to levels that no longer interfere with bacterial processes. Other resistance mechanisms involve changes in membrane permeability, particularly in gram-negative bacteria whose outer membranes act as selective barriers. By altering membrane channels or reducing the uptake of specific compounds, bacteria can limit the entry of antibiotics into the cell. 20
These adaptations illustrate an important reality of antimicrobial therapy: resistance is not simply the result of poor drug design or clinical misuse, although both can accelerate its spread. Instead, resistance emerges naturally whenever selective pressure favors organisms capable of surviving antimicrobial exposure. Understanding the biological mechanisms underlying resistance is therefore essential for developing new therapeutic strategies and preserving the effectiveness of existing drugs.

Expanding the Toolbox: Emerging Mechanisms
Although most existing antibiotics operate through a relatively small set of biological strategies, ongoing research continues to identify new ways to exploit bacterial vulnerabilities. Rather than focusing solely on incremental improvements to existing drug classes, many current discovery efforts aim to identify previously untapped targets or novel ways of interacting with known ones. These approaches do not replace established antibiotics, but they broaden the therapeutic toolbox and may help address pathogens that have developed resistance to older mechanisms.
One promising direction involves targeting the molecular precursors used to construct the bacterial cell wall. Teixobactin, a compound discovered through screening of previously uncultured soil microorganisms, illustrates this strategy. Instead of binding enzymes that assemble the wall, teixobactin attaches directly to lipid II and lipid III, essential building blocks of peptidoglycan synthesis. By sequestering these precursors, the drug prevents their incorporation into the growing cell wall, halting construction before it can begin.21,22 Because these lipid intermediates are highly conserved components of bacterial physiology, this mechanism may narrow the range of mutations that could readily confer resistance.
Another area of investigation focuses on the complex envelope surrounding Gram-negative bacteria. These organisms possess an outer membrane that provides an additional protective barrier and contributes to their resistance to many antibiotics. Several experimental compounds disrupt the machinery responsible for assembling this outer layer. Darobactin, for example, targets BamA, the central component of the β-barrel assembly machinery (BAM) complex that inserts proteins into the outer membrane. By interfering with this process, the drug prevents the insertion of outer-membrane proteins required for membrane integrity.23 Other candidates act on the systems that transport lipopolysaccharide molecules to the bacterial surface. Zosurabalpin has been shown to interfere with the LptB2FGC transporter that extracts lipopolysaccharide from the inner membrane during outer-membrane assembly.24 At the same time, murepavadin targets the related LptD component involved in lipopolysaccharide transport in P. aeruginosa.25 These strategies highlight how envelope biogenesis itself can serve as a drug target.
Novel mechanisms are also emerging even for targets that have been exploited for decades. Odilorhabdins show that a well-known structure, such as the ribosome, can still yield new opportunities for intervention. These peptides bind to the bacterial ribosome at a site not targeted by existing antibiotics, thereby altering translation accuracy and interfering with protein synthesis through a distinct mechanism.26,27
Finally, some experimental drugs target metabolic pathways that antibiotics have not traditionally exploited. Compounds that inhibit FabI, an enzyme involved in bacterial fatty acid biosynthesis, illustrate this approach. Because fatty acid production is essential for maintaining the bacterial membrane and other cellular structures, inhibiting this pathway can impair bacterial growth while potentially sparing beneficial microbes that rely on different metabolic systems.28
Conclusion — The Continuing Evolution of Antimicrobial Strategy
Despite the wide range of antibiotic molecules discovered over the past century, most successful antimicrobial drugs exploit a relatively small set of biological vulnerabilities. These strategies have proven remarkably effective because they target processes that bacteria cannot easily dispense with while leaving human cells largely unaffected.
At the same time, the evolutionary capacity of bacteria ensures that resistance will remain an ongoing challenge. Mutations that alter drug targets, enzymes that inactivate antibiotics, and cellular systems that reduce drug accumulation can all diminish the effectiveness of existing therapies. As these adaptations spread through microbial populations, antibiotics that once worked reliably may become less effective or require combination therapy to remain useful. Understanding the molecular basis of both antibiotic action and resistance, therefore, remains central to maintaining the effectiveness of antimicrobial treatments.
The continuing search for new antibiotics reflects this dynamic relationship between microbial evolution and drug discovery. Emerging compounds demonstrate that bacterial physiology still contains untapped vulnerabilities. Some experimental agents target components of cell-envelope assembly that were previously overlooked, while others bind familiar targets, such as the ribosome, in entirely new ways. Still others interfere with metabolic processes that earlier antibiotic classes have rarely exploited.
Antimicrobial strategy is not limited to the mechanisms discovered in the early decades of antibiotic research. Instead, it continues to evolve as researchers gain deeper insight into bacterial biology. Expanding the range of drug targets, improving stewardship of existing antibiotics, and developing therapies that reduce the likelihood of resistance will all play roles in sustaining the effectiveness of antimicrobial medicine in the years ahead.
In 2005, Chris Linaman was admitted to the hospital for surgery on a torn anterior cruciate ligament (ACL) in his left knee that he’d damaged playing basketball. Four harrowing months, four emergency surgeries, and $300,000 in medical costs later, he finally cleared the Methicillin-resistant Staphylococcus aureus (MRSA) infection he’d developed following the initial “routine” surgery.1
Antibiotics are some of the most powerful drugs ever developed, and have become the backbone of modern medicine; routine or major trauma surgery, cancer treatment, neonatal intensive care, and organ transplants all depend on antibiotics to prevent or treat infections that can occur when the body’s normal barriers or immune defenses are compromised. Without effective antibiotics, the risk of infection would make many of these interventions too dangerous to attempt. Yet they have become victims of their own success.
Drivers of Antimicrobial Resistance
Antimicrobial resistance (AMR) is the result of a naturally occurring evolutionary arms race, in which bacteria evolve to survive the drugs designed to kill them. Their ongoing changes render infections more difficult to treat, diseases easier to spread, and increase the risk of severe illness, disability, and death. Antimicrobial resistance is thought to have caused 1.27 million deaths in 2019 and contributed to 4.95 million.2 As the case study above illustrates, this is not only a problem in emerging economies but affects patients, hospitals, and health systems worldwide, regardless of geography or economic status. The economic impact of antimicrobial resistance is staggering; the World Bank estimates gross domestic product (GDP) losses of between 1 and 3.4 trillion US dollars by 2030 as a result of AMR, and over $1 trillion in additional healthcare costs by 2050.3
While bacterial evolution that enables survival of antibiotic treatment is a natural phenomenon, human intervention has significantly accelerated its pace. Poor sanitation, global travel, and underfunded surveillance schemes all contribute to the rising incidence of AMR. Still, it is the overprescription of antibiotics and the widespread agricultural use—or misuse—of antibiotics that have been the real drivers of this crisis.4
The Drug Development Paradox
At first glance, the market for antibiotics should be thriving due to AMR, given the enormous global patient population and demonstrable unmet need. And yet the opposite is true. There is next to no incentive for the private sector to invest in antibiotic development.
As recognition of the dangers of AMR has increased, so has responsible stewardship of antibiotics, creating a unique paradox within the pharmaceutical industry; the more successful a new antibiotic is, the less it will be prescribed.
Proper stewardship requires that antibiotics be used sparingly and for as short a period of time as possible. In fact, hospitals will actively hold back new antibiotics to be used as a “last line of defence” against a new resistant bacterial strain, significantly impacting sales of these medications.
Therefore, unlike almost any other class of drug, getting an antimicrobial product approved by the FDA is by no means a guarantee of commercial success. Achaogen, for example, filed for bankruptcy in 2019, less than a year after the FDA approved its first commercial antibiotic, Zemdri (plazomicin).5 This lack of commercial success is not an unusual circumstance, nor is it an “exception to a rule”. It is one of many similar stories and a stark explanation for why the number of potential antimicrobial treatments being developed by the world’s largest pharmaceutical companies has dropped by 35% in the last five years.6

Antibiotics as Global Infrastructure: A Top-Down Approach
When there is little incentive for the private sector to address AMR, governments must step in. The Global Fund to Fight AIDS, Tuberculosis, and Malaria, the Coalition for Epidemic Preparedness Innovations, and the US National Cancer Institute Moonshot Initiative set a precedent for governments intervening when the stakes are high and demonstrate that such interventions can reshape biomedical innovation.
In fact, the Organisation for Economic Co-operation and Development (OECD), an intergovernmental forum of 38, mostly developed, member countries that promotes policies to improve economic and social well-being worldwide, has already resolved that investing in effective, cost-effective policies will be key to tackling AMR.7 Given that the root causes of AMR are misuse of antibiotics in both humans and animals, the OECD further concludes that fighting AMR will require a One Health approach: coordinated policy action across human, animal, and plant health, agri-food systems, and the environment.
Initiatives such as this transition antibiotics from “private assets” to public goods. How to fund and maintain equitable access to these lifesaving drugs then becomes a societal challenge.
Several policy initiatives have been proposed to address this, including both “push” and “pull” funding techniques to re-incentivise drug development and commercialisation, regulatory reform to ease the path of new antibiotics to market, and global surveillance schemes to monitor the incidence of AMR, track the evolution of new resistant strains, and coordinate a global response.
“Push” funding initiatives
Push funding is a mechanism to revitalize the antimicrobial drug pipeline by supporting the discovery and development of new therapeutics. Notable funding schemes include:
- CARB-X: One of the largest antibiotic R&D accelerators, funded by both the US and UK Governments as well as charitable foundations. CARB-X funds research into novel antibiotics, vaccines, and diagnostics for AMR and has, to date, invested over $506 million in 121 research projects, of which 23 have entered clinical trials, and three are on the market. Notably, while CARB-X funding is non-dilutive, it requires innovators to have already secured some private funding for their projects.8
- GARDP: The Global Antibiotic Research and Development Partnership is a nonprofit organization that funds the development of antibiotics for neglected infections through a public-private partnership model and prioritizes public health impact, affordability, and high-burden countries.9
- The AMR Action Fund: A private-public venture capital fund launched in 2020 by major pharmaceutical companies and global health organizations to support antimicrobial resistance. The AMR Action Fund’s goal is to bring two to four new antibiotics to market by 2030.10
However, while these programs help build a healthy pipeline of antimicrobial assets, they are not sufficient to ensure the commercial success and, therefore, the longevity of these products once they’re approved. This is where “pull” funding comes in.
“Pull” funding initiatives
Pull funding schemes, such as subscription models, aim to reincentivize the commercialization of new antibiotics by decoupling revenue from sales volume and basing payment on the drug’s value to public health. In this type of model, governments pay companies a fixed annual fee for access to an antibiotic, allowing companies to maintain a predictable revenue stream. In contrast, the antibiotics themselves can still be carefully stewarded and held in reserve for resistant infections.
The UK’s National Health Service piloted the first such subscription model in 2022, signing multi-million-pound contracts per year for the antibiotics ceftazidime-avibactum and cefiderocol, regardless of how much of each drug is used. Following this pilot scheme, the antibiotic subscription model was established as a formal commissioning route for new antimicrobial drugs in the UK, with the first round of procurement occurring in August 2024.11,12
Regulatory reform
Regulatory reform is another key pillar of a top-down solution to antimicrobial resistance, since small patient populations for emerging infections and ethical implications around placebo treatments make traditional clinical trials difficult and expensive for novel antimicrobials.
The US LPAD Pathway was introduced in 2016 and is designed to accelerate approvals for drugs intended to treat serious or life-threatening infections in limited patient populations with unmet needs. These drugs may be approved based on smaller, shorter, or fewer clinical trials, reducing the cost, complexity, and timeline of bringing a new antibiotic to market.13
International surveillance
The final pillar of a top-down solution discussed in this article is international surveillance that tracks the emergence of resistant pathogens and antibiotic use in a standardized way, enabling national data to be aggregated into global datasets. This is exactly what the World Health Organization’s Global Antimicrobial Resistance and Use Surveillance System (GLASS) initiative was established to achieve. The trends that initiatives such as this help to detect can then inform future research. For example, in their 2025 report, GLASS found that resistance to antibiotics currently used to treat the most severe infections is increasing among some Gram-negative pathogens14, which in turn is increasing reliance on last-resort treatments. The identification of such trends is an essential first step to countering them.
Collectively, these initiatives demonstrate how seriously global authorities are taking the threat of AMR, and offer significant hope for the fight against it. However, to make a sustained impact over the long term, push-and-pull funding, regulatory reform, and international surveillance must be implemented globally and in tandem to ensure that the world maintains access to a steady supply of effective antibiotics.

The Bottom-Up Countermeasures: Reducing the Need for Antibiotics
The adage “a prevention is better than a cure” is never truer than in the case of antimicrobial infections, where reducing bacterial infections is the ultimate route to reduced antibiotic use and, therefore, bacterial resistance.
Endeavors to prevent bacterial infections fall into three major categories: improved sanitation, prevention of hospital-acquired infections, and reduced antibiotic use in agriculture and food production. And while, in every case, the ultimate success of the initiative requires grassroots adoption (better handwashing, routine cleaning, and improved animal husbandry on farms around the world, for example), the mandate, implementation, and maintenance of schemes to enforce these practices still require top-down infrastructure and policy shifts.
Sanitation and clean water
The World Health Organization identifies water, sanitation, and hygiene (WASH) as a central bottom-up approach to reducing AMR. Poor sanitation leads to diarrheal diseases, typhoid, cholera, and other intestinal infections, which in turn result in high levels of antibiotic use. Resistant bacteria enter the environment through untreated sewage, hospital wastewater, and agricultural run-off, facilitating the spread of resistance genes between bacteria and, without safe water systems, returning to humans and propagating antibiotic-resistant infections among communities.
Resistant bacteria don’t just spread in hospitals; they travel through contaminated water. Clean drinking water and proper sewage treatment are among the most effective tools in the AMR prevention toolkit. At the same time, grassroots initiatives like handwashing campaigns can be highly successful in reducing disease transmission. Indeed, the Centers for Disease Control and Prevention (CDC) notes that handwashing education can reduce the number of people suffering from diarrheal infections by 23-40%.15
Preventing hospital infections
The very nature of hospitals, with high levels of antibiotic use, and the number of vulnerable patients and invasive procedures being undertaken, makes these extremely high-risk environments for the emergence and spread of AMR. In the US alone, at least one patient in every 31 has a healthcare-associated infection on any given day.16
Again, hand hygiene and strict cleaning protocols, not just for surgical instruments, but for any high-touch surfaces and patient rooms or waiting areas, are key to preventing infection. More complex initiatives include screening high-risk patients (such as those hospitalized at home after initial treatment abroad), rapid diagnosis of resistant bacterial infections, strict isolation of affected patients, and ongoing stewardship of antibiotic prescribing.
Reducing antibiotic use in agriculture and food production
When livestock are farmed intensively, often in crowded and unsanitary conditions, antibiotics are often given as a preventive measure to prevent infections from developing due to poor husbandry practices. Farm animals consume about 30% of all antibiotics in the UK and 65% worldwide. In the UK, intensively reared pigs and poultry account for 79% of antibiotic use.17 This is a major driver of AMR, and resistant bacteria can then spread from animals to humans through contaminated meat, from farm workers, or via environmental run-off into water systems.
Reducing the level of antibiotic use in farming requires both top-down policy changes, such as the 2006 EU ban on antibiotic growth promoters in livestock18, and the provision of guidelines to support veterinary stewardship, and grassroots implementation of better farming conditions, including reducing the density of animals in any given space, improving hygiene, and implementing vaccination programs for farm animals.
The most effective tools against antimicrobial resistance are, on the surface, very straightforward: clean water, safer hospitals, and food systems that rely less on routine antibiotic use. These sound like simple solutions, but implementation and global adoption are anything but.
The problem is Clear. The Tools to Reduce AMR Exist. What Now?
While it has been widely described as a slow pandemic, AMR is perhaps more accurately equated to climate change than to an emerging pathogen. The problem is known, and the scale is enormous. However, solutions have been defined, and the tools to address them largely exist. The challenge now is the will to act and implement. How to align economic incentives with public health infrastructure and global cooperation? The danger of AMR is not that we lack the science to confront it. It’s that we have yet to implement the systems required to wield that science effectively.
Mapping the antimicrobial resistance (AMR) landscape requires moving beyond the traditional view of drug development as a linear pipeline and instead understanding it as an interconnected, interdependent system. Each stage, from discovery through stewardship, is tightly coupled, with weaknesses in one segment reverberating across the entire ecosystem.
At the front end, discovery is driven by a network of academic institutions, public funders, and early-stage biotech innovators. Universities such as the University of Oxford, Harvard University, and Rockefeller University remain foundational, combining deep expertise in microbiology, infectious disease, and structural biology with access to advanced research infrastructure. Their work, often curiosity-driven, generates the mechanistic insights and early validation that seed therapeutic development.
This activity is sustained by public and philanthropic funding bodies, including the National Institutes of Health, the Wellcome Trust, the Innovative Medicines Initiative, and others, which provide the non-dilutive capital required to advance early-stage research that would otherwise lack commercial backing. Together, these institutions form a discovery engine that is scientifically productive but economically fragile, particularly in areas such as gram-negative pathogens, where technical complexity and low expected returns have driven many large pharmaceutical companies out of the field.¹
Early-stage biotech companies act as the translational bridge between discovery and development. Firms such as VenatoRx Pharmaceuticals, Nosopharm, Locus Biosciences, and Pherecydes Pharma are advancing novel modalities, from next-generation β-lactamase inhibitors to CRISPR-based antimicrobials and bacteriophage therapies. Yet their ability to progress candidates depends heavily on downstream system integrity.
That dependency becomes most apparent in the translational gap, often referred to as the “valley of death.” Here, organizations such as CARB-X, the Global Antibiotic Research and Development Partnership (GARDP), and the Biomedical Advanced Research and Development Authority (BARDA) play a critical role in advancing candidates through preclinical and early clinical development. These entities provide both funding and technical support, helping to bridge the gap between scientific promise and later-phase clinical studies.² Despite these efforts, this stage remains defined by a severe funding cliff. Advancing a candidate through IND-enabling studies requires millions of dollars, yet access to venture capital remains constrained by weak commercial prospects.11 As a result, many promising programs stall before reaching clinical development.
Clinical Development Bottlenecks
Clinical development presents a distinct set of challenges that differ markedly from those in other therapeutic areas. This stage is led by biotech sponsors, including companies such as Entasis Therapeutics, Venatorx Pharmaceuticals, Iterum Therapeutics, Nabriva Therapeutics, Spero Therapeutics, and Forge Therapeutics, often operating with limited capital while navigating complex regulatory and operational demands.
These sponsors rely heavily on contract research organizations (CROs) such as IQVIA, Parexel, PPD, ICON plc, and Medpace, which provide global trial infrastructure, alongside specialized contributors like Certara for pharmacokinetic and pharmacodynamic modeling. Clinical trial execution is further supported by networks such as the Antibacterial Resistance Leadership Group (ARLG), the European Clinical Research Alliance on Infectious Diseases, and the UK’s National Institute for Health and Care Research Clinical Research Network.
Yet even with this infrastructure, AMR trials remain fundamentally misaligned with traditional clinical models. Resistant infections are sporadic and geographically dispersed, making patient recruitment difficult. Trial designs, often centered on non-inferiority frameworks, can be ill-suited to situations in which existing therapies are no longer effective.³
These challenges underscore a broader insight: clinical development paradigms designed for chronic, high-prevalence diseases are often ill-suited to AMR. Emerging approaches, including platform trials, adaptive designs, and globally coordinated trial networks, offer a path forward but remain underutilized.
Manufacturing and Supply Resilience
Once an antimicrobial candidate succeeds clinically, its impact depends on the resilience of a highly distributed manufacturing ecosystem. This network includes CDMOs such as Recipharm, Lonza, Catalent, Almac Group, and Delpharm, alongside manufacturers like Hikma Pharmaceuticals that operate with CDMO-like capabilities in injectables.
Upstream, API production is concentrated among players such as ACS Dobfar, Sandoz, Teva Pharmaceutical Industries, Aurobindo Pharma, Zhejiang Hisun Pharmaceutical, and North China Pharmaceutical Group. These companies form the backbone of global antibiotic supply, yet their geographic concentration introduces systemic risk. A significant proportion of antibiotic APIs are produced in China and India, creating vulnerability to geopolitical disruption, regulatory shifts, and environmental constraints.⁴
Economic realities compound this risk. Antibiotics are typically low-margin, low-volume products, making them less attractive than oncology, rare disease, or biologics programs. As a result, manufacturers must continuously balance capacity allocation, often deprioritizing antimicrobial production.
The COVID-19 pandemic exposed the fragility of this system, with disruptions to raw materials and logistics triggering shortages across multiple drug classes.² These dynamics reinforce a critical point: even clinically successful antibiotics can fail to reach patients if manufacturing systems are not economically sustainable.
Market Failure and Commercial Exit
At commercialization, the AMR ecosystem faces one of its most well-documented failures: a market that fails to reward innovation.
Recent history provides multiple examples. Companies such as Achaogen and Melinta Therapeutics entered bankruptcy shortly after bringing new antibiotics to market, highlighting a structural disconnect between regulatory approval and commercial viability.⁵
This paradox reflects a fundamental misalignment. Antibiotics are designed to be used sparingly to preserve efficacy, yet traditional pharmaceutical business models are volume-based. Emerging policy approaches seek to address this issue. “Delinkage” models propose separating revenue from sales volume, while subscription-based systems, such as those piloted in the United Kingdom and proposed in the United States through the PASTEUR Act, provide fixed payments for access to critical antibiotics.⁶
Without such reforms, the antibiotic pipeline will remain unstable, not due to a lack of innovation, but because the system and revenue models are not structured properly to sustain it.
Stewardship, Access, and Distribution
The final stage of the AMR supply chain determines how antimicrobials are used in practice. Hospitals, governments, NGOs, and global health organizations are responsible for balancing access with appropriate use.
This creates a persistent tension. In some regions, antibiotics are overused due to limited regulation and diagnostic capacity. In others, access remains constrained, with essential medicines unavailable or unaffordable. The World Health Organization has established global stewardship frameworks emphasizing optimized prescribing, surveillance, and education, yet implementation remains uneven.¹
Diagnostics and Surveillance
Diagnostics and surveillance function as the information backbone of the AMR system. Diagnostic companies, clinical laboratories, and public health agencies generate and analyze data that inform treatment decisions and track resistance patterns.
Despite their importance, these systems remain underdeveloped. Many healthcare settings still rely on empirical treatment due to a lack of rapid diagnostics, contributing to inappropriate antibiotic use and accelerating resistance.³
The Missing Link: Financing and Incentive Alignment
Across every node of the AMR supply chain, a common theme emerges: misaligned incentives.
Push funding mechanisms, including grants and early-stage investments from organizations such as CARB-X and BARDA, have revitalized discovery and supported translational progress. However, without complementary pull incentives, products struggle to achieve commercial sustainability.
The result is a fragmented system in which each stakeholder optimizes for its own mandate, whether scientific innovation, financial return, or public health outcomes, but the system as a whole fails to deliver consistent impact.
Integration: From Fragmentation to Function
The AMR supply chain today is defined not by a lack of innovation, but by a lack of coordination.
Public–private partnerships, including CARB-X and GARDP, demonstrate how stakeholders can be aligned across academia, industry, and government. Expanding these models, alongside shared data platforms and end-to-end funding frameworks, will be essential to improving system performance.
The defining insight is clear. A single breakthrough molecule will not solve AMR. It will be solved, or not, by whether we can build a system capable of consistently translating scientific potential into durable clinical impact.
Antibiotic Resistance Continues to Outpace Drug Development
The number of bacterial pathogens capable of resisting multiple antibiotics continues to rise, including resistance to some recently introduced drugs.1,2 This growing challenge is reflected in the World Health Organization’s (WHO) priority pathogen list, which identifies organisms for which urgently needed new therapies remain scarce.3 At the top of this list are several gram-negative bacteria — Acinetobacter baumannii, Pseudomonas aeruginosa, and members of the Enterobacterales family — along with major gram-positive threats, such as Staphylococcus aureus, Enterococcus spp., and Streptococcus pneumoniae. Collectively, these organisms account for a large share of difficult-to-treat infections in both hospital and community settings.
The problem extends beyond classical genetic resistance. Many pathogenic bacteria acquire mutations that allow them to survive exposure to antibiotic molecules that would previously have been lethal. Other populations display antibiotic tolerance, remaining viable (though not actively proliferating) during transient drug exposure. Still others form persistent subpopulations that enter dormant states capable of surviving even very high antibiotic concentrations.4 These multiple survival strategies allow pathogens to withstand treatment, recover after therapy ends, and ultimately contribute to the continued expansion of antimicrobial resistance.
As a result, even as new antibiotics enter the market, many of them remain vulnerable to the same biological defenses that undermine older drugs. Addressing antimicrobial resistance will therefore require approaches that extend beyond traditional antibiotic mechanisms and draw on a broader therapeutic toolkit.

Moving Beyond the Classical Antibiotic Paradigm
Analyses published in early 2020 by both the WHO and the Pew Charitable Trusts reached a sobering conclusion: none of the antibiotics currently available worldwide can reliably address many clinically significant drug-resistant bacterial infections.4 While some new antibiotics have been introduced in recent years, most represent incremental improvements rather than fundamentally new solutions. Between the publication of the WHO priority pathogen list in 2017 and late summer 2025, the U.S. Food and Drug Administration (FDA) approved 16 antibiotics. Yet, nearly all were from previously established antimicrobial classes.1 As a result, many remain vulnerable to resistance mechanisms that bacteria have already evolved.
Traditional antibiotics operate through a relatively limited set of biological mechanisms. Most act by inhibiting bacterial cell wall synthesis, disrupting cell membrane function, or interfering with protein synthesis by binding to ribosomal subunits.5 Bacteria can evade these effects through several well-established strategies, including reducing drug uptake, accelerating drug metabolism, enzymatically inactivating antibiotics, or modifying the molecular targets to which the drugs bind.4,6 Once these mechanisms emerge and spread, new drugs that rely on the same underlying modes of action often face the same fate.
Expanding the antimicrobial arsenal will therefore require therapies that operate outside the traditional antibiotic paradigm and bypass these entrenched resistance pathways. A growing body of research is exploring alternative therapeutic modalities that target bacteria through fundamentally different mechanisms. Prominent examples include bacteriophage-based therapies, antibody-mediated antibacterial strategies, microbiome-derived treatments, host-directed therapies that strengthen immune defenses, and synthetic polymeric antibiotics.
Harnessing Bacteriophages to Combat Drug-Resistant Bacteria
Bacteriophage therapy is among the most conceptually distinct alternatives to traditional antibiotics. Bacteriophages, a class of viruses that infect and kill bacteria, target specific bacterial species and replicate within them, creating a self-amplifying antimicrobial effect. Each phage attaches to a bacterial cell and introduces its genetic material, initiating one of two possible biological cycles. In the lytic cycle, the phage takes over the host cell’s machinery to produce new phage particles, ultimately causing the bacterial cell to rupture and release additional viruses. In the lysogenic cycle, phage DNA integrates into the bacterial chromosome or persists in the cytoplasm as an episome, becoming a prophage that is replicated as the bacterium divides and can later transition into the lytic cycle under environmental stress.7
Because bacteriophages infect bacteria rather than human cells, they do not damage host tissues and generally do not disrupt the broader human microbiome like broad-spectrum antibiotics.8 Their high specificity also allows targeted elimination of pathogenic bacteria while leaving beneficial microbial communities largely intact. Another advantage lies in their evolutionary adaptability: although bacteria can develop resistance to individual phages, phages themselves can evolve in response to bacterial defenses, creating a dynamic biological arms race that differs fundamentally from static chemical antibiotics.
The therapeutic use of bacteriophages is not a new concept. Phage therapy was first explored more than a century ago and continues to be used in parts of Eastern Europe as a treatment option when conventional antibiotics fail.7 Modern research has expanded on this foundation, demonstrating that phage therapy often performs best when multiple phages are combined into phage cocktails, which broaden the range of bacterial targets and reduce the likelihood that resistance will emerge.
Recent preclinical studies involving pathogens, such as P. aeruginosa, A. baumannii, and Klebsiella pneumoniae, have shown promising results.9 Early clinical experience, largely derived from compassionate-use cases, suggests that combining phages with conventional antibiotics may further enhance effectiveness in patients with multidrug-resistant infections. These phage–antibiotic combinations (PACs) have been reported to enhance bacterial killing, suppress the emergence of resistance, and, in some cases, restore susceptibility to antibiotics that had previously lost efficacy.10
Despite these advances, important practical challenges remain. Many phage therapies must be tailored to the specific bacterial strain causing a patient’s infection, requiring rapid identification of the pathogen and testing candidate phages against it.9,10 Standardized clinical workflows for evaluating patient isolates are still limited, and the time required for strain identification and phage matching can restrict the use of personalized phage therapy in rapidly progressing infections.
To address these limitations, researchers are developing off-the-shelf phage cocktails designed to cover broader bacterial populations. One recent study employed a screening strategy to identify “complementarity groups” of phages targeting different bacterial receptors and to examine their interactions with specific antibiotic classes. The resulting phage–antibiotic combinations demonstrated high efficacy against P. aeruginosa and S. aureus individually and when used together against mixed bacterial cultures.11
Although these findings highlight the potential of phage therapy as a powerful addition to the antimicrobial toolkit, the field still lacks large, systematically designed clinical trials. Existing studies involve diverse phage–antibiotic combinations administered through varying routes, dosing schedules, and treatment durations, making it difficult to draw definitive conclusions about optimal therapeutic strategies.10 Continued clinical investigation will be necessary to determine how phage-based therapies can be integrated most effectively into the treatment of drug-resistant infections.
Importantly, phage therapy illustrates how expanding beyond conventional antibiotics can introduce entirely different biological strategies for controlling bacterial infections: approaches that exploit viral replication and evolutionary dynamics rather than relying solely on static chemical inhibition.
Antibody Therapies: Precision Targeting of Bacterial Pathogens
Monoclonal antibodies (mAbs) represent another strategy for expanding the antimicrobial toolkit beyond conventional antibiotics. Traditional small-molecule antibiotics, while effective in many settings, often act broadly against bacteria and can disrupt beneficial microbial communities as well as pathogens. They may also persist in the environment and exert selective pressure that accelerates the development of resistance. These limitations have prompted growing interest in antibody-based approaches that target bacterial pathogens with far greater precision.12,13
Antibodies can be engineered to recognize highly specific bacterial antigens, allowing them to attack pathogenic organisms while minimizing collateral damage to the surrounding microbiome. Their pharmacokinetic properties also offer practical advantages: many antibodies exhibit relatively long half-lives in circulation, which can reduce dosing frequency and potentially improve treatment adherence. Because they are produced via cell culture rather than chemical synthesis and degrade naturally in biological environments, antibody therapeutics may also offer environmental advantages over traditional antibiotics.
Unlike small molecules, which typically disrupt bacterial physiology directly, antimicrobial antibodies act through a range of immune-mediated mechanisms. These include neutralizing bacterial toxins, blocking receptor-mediated bacterial adhesion to host tissues, interfering with biofilm formation, promoting immune clearance through antibody-mediated complement killing, and enhancing opsonophagocytosis (antibody tagging of pathogens that facilitates their engulfment by phagocytic immune cells). Antibodies can also bind outer-surface proteins associated with antibiotic resistance, thereby weakening bacterial defenses. By mobilizing multiple immune mechanisms rather than relying on a single biochemical target, antibody-based therapies may reduce the likelihood that bacteria will rapidly develop resistance.12,13
The versatility of antibody engineering further expands their therapeutic potential. Small molecule antibiotics can be conjugated to antibodies to create antibody–antibiotic conjugates, enabling targeted delivery of antibacterial payloads directly to infected cells while limiting off-target bacterial killing. Bispecific antibodies can simultaneously bind two different bacterial antigens or epitopes, creating a coordinated multipoint attack. Researchers are also exploring alternative antibody formats, including nanobodies and single-chain antibody fragments (scFvs), which may offer improved tissue penetration or manufacturing advantages.
Several antibody therapies targeting bacterial pathogens have already reached the market. Examples include raxibacumab (GlaxoSmithKline) and obiltoxaximab (Elusys Therapeutics), which neutralize Bacillus anthracis toxins in bioterrorism preparedness applications, and bezlotoxumab (Zinplava, Merck & Co.), which helps prevent recurrence of Clostridioides difficile infection in high-risk patients.12
Despite these advances, clinical development of antibacterial antibodies has proven challenging. Only a limited number of candidates have progressed into late-stage trials, and many programs have stalled during phase I or phase II studies.13 Early efforts often focused on fully humanized antibodies targeting pathogens, such as S. aureus, P. aeruginosa, or Clostridium botulinum. Several factors contributed to their limited clinical success. Bacterial infections frequently involve multiple pathogens, antigenic variability can reduce antibody effectiveness, and conventional monoclonal antibodies often rely on a single mechanism of action. For example, antibodies that neutralize bacterial toxins may limit tissue damage after infection has begun but may not directly eliminate the underlying bacteria.
Newer strategies aim to address these limitations by expanding the functional capabilities of antibody therapeutics. Antibody–antibiotic conjugates combine the targeting precision of antibodies with the bactericidal activity of conventional antibiotics, while bispecific antibodies can simultaneously target multiple bacterial antigens. Researchers are also investigating antibody cocktails that include several antibodies targeting different targets, potentially broadening their activity against diverse pathogens and reducing the likelihood of resistance.13
Microbiome Therapeutics: Restoring the Body’s Natural Defenses
The human microbiome plays a central role in maintaining resistance to infection, and disruption of this complex microbial ecosystem can significantly increase vulnerability to pathogenic bacteria. Such disruption can lead to dysbiosis, defined as an imbalance in the composition and functional capacity of microbial communities.14 Dysbiosis has been associated with numerous disease states,15 and the gut microbiota also plays an important role in drug metabolism and host immune regulation.14 When the composition or activity of the microbiome changes, susceptibility to infection can increase as protective microbial functions are weakened.16
These microbial communities influence host biology through multiple mechanisms. Alterations in microbiome composition can affect the production of antimicrobial peptides, influence epigenetic regulation and gene expression patterns, modify enzymatic activity through microbiome-derived metabolites, and shape inflammatory and cellular immune responses. Changes in microbial populations can also alter pH levels, influence nutrient availability for pathogens, and modify drug pharmacokinetics, including the activity and effectiveness of antibiotics.16 Through these interconnected pathways, microbiome disruptions can create conditions that allow pathogenic bacteria to establish infections more easily.
Microbiome-based therapeutics aim to restore or reshape these microbial ecosystems rather than directly killing pathogens. By promoting beneficial microbial populations, these therapies can help reestablish colonization resistance and reduce susceptibility to infection without the broad-spectrum bacterial killing often associated with conventional antibiotics.4 Several classes of microbiome-directed therapies are under development, including prebiotics, which serve as substrates for beneficial microbes; probiotics, which introduce live microorganisms with defined biological functions; synbiotics, which combine prebiotics and probiotics; and postbiotics, which comprise microbial metabolites or cellular components that influence microbiome activity.14
Probiotics in particular have attracted significant interest as researchers and biotechnology companies explore the use of defined microbial consortia — often referred to as live biotherapeutic products (LBPs) — to treat infectious and inflammatory diseases.17 Candidate organisms under investigation include Akkermansia muciniphila, Faecalibacterium prausnitzii, Veillonella, Ruminococcus, Christensenella minuta, and Bacteroides fragilis.14 Advances in genetic engineering are also enabling the design of modified microbial strains with enhanced targeting capabilities or therapeutic functions.16
One of the clearest demonstrations of microbiome-based therapy involves the treatment of recurrent or multidrug-resistant C. difficile infection (CDI). In these cases, fecal microbiota transplantation (FMT) from healthy donors has proven highly effective in restoring microbiome diversity and preventing recolonization by C. difficile spores. A donor-derived microbiome therapy, Rebyota (Ferring Pharmaceuticals), has received FDA approval for this indication.14 More recently, oral capsule formulations containing lyophilized microbiota have been developed, offering a simpler and less invasive treatment option. VOWST (Seres Therapeutics) is the first FDA-approved product of this type. Researchers are also evaluating microbiome transplantation approaches for other multidrug-resistant gastrointestinal infections and recurrent bacterial vaginal infections.14
Additional strategies under investigation include microbiome-based vaccines and combination approaches in which microbiome therapeutics are administered alongside conventional antibiotics to enhance treatment outcomes.16 By reinforcing the body’s natural microbial defenses, these approaches illustrate another pathway for expanding antimicrobial strategies beyond traditional antibiotic mechanisms.
Turning the Host into the Antimicrobial Target
Another strategy for addressing antimicrobial resistance focuses on strengthening the host rather than directly attacking bacterial pathogens. Host-directed therapies (HDTs) seek to enhance innate or adaptive immune responses or modulate cellular pathways that pathogens rely on for survival and replication.18,19 Because these approaches target host processes rather than bacterial structures, pathogens may find it more difficult to evolve resistance. HDTs interfere with intracellular and intercellular pathways that bacteria exploit during infection, thereby disrupting the conditions required for pathogen persistence.18,20
Several host-directed strategies have been explored in the treatment of chronic infectious diseases, such as tuberculosis. Many of these therapies focus on regulating inflammatory signaling pathways. Agents targeting cytokine activity, including antibodies directed against interleukin-6 or tumor necrosis factor, have been investigated alongside compounds such as interferons, antioxidants like N-acetylcysteine, vitamins including vitamin D, and anti-inflammatory drugs, such as statins and cyclooxygenase-2 inhibitors.18,20 By modulating immune signaling, these therapies aim to restore effective host responses while limiting excessive inflammation that can damage tissues.
Other HDTs work by stimulating innate antimicrobial mechanisms within host cells. Research has explored compounds that increase production of antimicrobial peptides (AMPs), reactive oxygen species (ROS), and nitric oxide (NO), as well as therapies that induce autophagy, a cellular process that degrades intracellular pathogens.21 These molecules and pathways form part of the body’s natural antimicrobial defenses and contribute to the barrier against microbial invasion. Many multidrug-resistant pathogens suppress these protective mechanisms during infection. Therapies that stimulate their production can therefore help restore the host’s ability to eliminate invading bacteria. In some cases, these mechanisms may act synergistically when activated together.
Epigenetic regulators provide another route for activating antimicrobial defenses. Histone deacetylase inhibitors (HDACis), such as butyrate and other short-chain fatty acids (SCFAs), can increase the production of antimicrobial peptides while strengthening epithelial barrier function by promoting tighter cell junctions.21 However, direct administration of butyrate can trigger undesirable immunogenic reactions in the gastrointestinal tract, limiting its clinical utility. Alternative molecules such as phenylbutyrate have demonstrated promising activity in animal models. Vitamin D has also attracted interest because it promotes autophagy and can enhance antimicrobial activity when combined with other host-directed therapies.
Several classes of drugs already approved for other diseases may also be repurposed as host-directed antimicrobial therapies. Immunomodulatory drugs, certain cellular and gene therapies, and antibody-based treatments originally developed for cancer are now being evaluated for infectious diseases.19 Current research focuses heavily on chronic infections, such as tuberculosis, hepatitis B and C, and HIV-1, but similar strategies may prove applicable to a broader range of bacterial pathogens.
Despite their promise, host-directed therapies present important challenges. Because these interventions alter host cellular pathways, they carry the risk of unintended toxicities.19 Their effectiveness may also depend on careful timing relative to infection stage and disease progression. Successful HDT development, therefore, requires identifying host pathways that pathogens depend upon but that can be safely modulated without disrupting essential physiological functions.
Programmable Polymers for Combating Drug-Resistant Bacteria
Synthetic antimicrobial polymers (SAPs) represent another emerging strategy for combating drug-resistant infections. Unlike conventional antibiotics that typically interfere with specific intracellular targets, many SAPs act directly on bacterial membranes. Their antimicrobial activity derives in part from their relatively large molecular size and in part from the ability to engineer their chemical structure by selecting different polymer backbones and functional substituents.22 Because most polymers cannot readily penetrate bacterial cells, they instead interact with the negatively charged outer surfaces of both gram-positive and gram-negative bacteria. Positively charged polymers, including poly(quaternary ammonium) compounds, polyethylenimine, and polyamines, can bind to these surfaces and disrupt membrane integrity, leading to bacterial cell death through mechanisms that are inherently difficult for pathogens to evade through conventional resistance pathways.22
Researchers have explored a wide range of SAP architectures. Some polymers exhibit intrinsic antimicrobial activity, while others carry attached bioactive functional groups or act as delivery vehicles for antibacterial agents.22 Most SAPs are cationic, but anionic and amphiphilic polymers containing both positive and negative charges have also been investigated. Functional groups incorporated into polymer backbones often include cationic, hydrophobic, π-electron–containing, amphiphilic, and metal-binding moieties, each of which can influence antimicrobial activity. In addition, stimuli-responsive polymers are being designed to activate under specific environmental conditions, such as changes in pH or temperature, or exposure to oxidative environments or enzymes, enabling more targeted antimicrobial effects.
Many SAPs have already been applied as antimicrobial coatings for medical devices and implants to reduce contamination and infection risk.22 Increasingly, however, these materials are being investigated as systemic antimicrobial therapies. Experimental polymers have demonstrated activity against pathogens such as Escherichia coli and Salmonella, including multidrug-resistant strains.
A closely related class of materials — synthetic antimicrobial peptides (AMPs) — has also attracted significant interest. Natural AMPs are key components of innate immune defenses, and synthetic variants can be engineered to disrupt bacterial membranes while also influencing immune and inflammatory responses.23 The development of clinical AMP candidates has proven challenging due to issues such as toxicity, limited in vivo stability, and manufacturing complexity. Advances in computational design are helping address these limitations. Deep learning approaches are now being used to identify peptide sequences with improved antimicrobial activity and safety profiles.
Machine learning is also accelerating the broader design of antimicrobial polymers. In one recent example, researchers identified three poly(β-amino ester) polymers with strong activity against drug-resistant bacteria, low toxicity, and the ability to self-assemble into stable nanoparticles (SANPs).23 When combined with penicillin G, these polymers restored susceptibility in drug-resistant gram-positive bacteria, illustrating how polymeric materials can enhance the effectiveness of existing antibiotics.
Other approaches combine polymer chemistry with traditional antibiotics to overcome specific resistance mechanisms. One self-assembling nanocomplex composed of a guanidinium-functionalized cationic polymer and the anionic prodrug colistin methane sulfonate was developed to target bacteria carrying plasmid-borne mobilized colistin resistance (mcr) genes.24 By delivering both the antibiotic and the antimicrobial polymer simultaneously, this system was able to reverse colistin resistance and effectively kill mcr-positive strains of multidrug-resistant E. coli and K. pneumoniae.
Polyacrylamide-based copolymers represent another promising platform. These materials disrupt bacterial membranes while maintaining selectivity for bacterial cells over mammalian cells and have demonstrated efficacy against several multidrug-resistant pathogens.25 When administered alongside conventional antibiotics, these SAPs can also enhance antibiotic activity against resistant bacteria.
These polyacrylamide polymers offer several practical advantages, including commercial availability, chemical stability, and relatively straightforward synthesis.25 Effective candidates typically incorporate a cationic monomer to support membrane binding, a hydrophilic monomer to maintain water solubility, and a smaller fraction of hydrophobic monomer designed to promote membrane disruption. Leading candidates have demonstrated activity against pathogens such as Staphylococcus aureus, E. coli, K. pneumoniae ATCC 13884 (gram-negative), and Enterococcus faecium ATCC 35667 (gram-positive). Notably, four of these polymers showed activity against all four pathogens.
Not all antimicrobial polymers are large macromolecules. Researchers have also developed shorter, biodegradable oligomeric polymers capable of penetrating bacterial membranes. One example involves poly(imidazolium ester) polymers that cross bacterial membranes and interact directly with nucleic acids.26 In vitro studies showed that these polymers bind DNA through electrostatic and hydrogen-bond interactions to form biomolecular condensates. This phase separation disrupts transcription and translation processes and may also interfere with RNA synthesis. In preclinical animal models, the leading candidate safely treated systemic, intramuscular, and pulmonary infections caused by drug-resistant bacteria and fungi, including S. aureus, S. epidermidis, E. coli, K. pneumoniae, and Candida albicans, at relatively low doses.
By enabling antimicrobial activity through engineered membrane disruption, programmable nanostructures, and novel intracellular mechanisms, polymer-based therapeutics demonstrate how materials science and molecular engineering can broaden the range of strategies for combating antimicrobial resistance.

Rethinking Antibiotic Development for the AMR Era
Scientific innovation alone will not resolve the antimicrobial resistance crisis if economic and development incentives remain misaligned. One of the most persistent barriers to progress is the limited investment in antibiotic development by major pharmaceutical companies. According to one recent analysis, only six large biopharma companies currently maintain small antibiotic development programs, while several biotechnology firms focused on antibiotic innovation have entered bankruptcy.27 This contraction reflects a difficult economic reality: new antibiotics are often reserved for the most severe or resistant infections to preserve their effectiveness, which significantly limits the revenue they can generate.
These economic constraints have prompted some researchers to question whether the traditional strategy of developing highly targeted antibiotics for individual resistant pathogens is sustainable. Treatments designed to address specific multidrug-resistant organisms often serve relatively small patient populations, and the growing diversity of resistant bacteria would require developing many separate drugs to address each threat.27 As a result, some investigators have proposed a shift toward “pathogen-general” antimicrobial approaches that could work across a wider range of bacterial species.
One proposed solution involves the development of general potentiators — agents that enhance the effectiveness of existing antibiotics against both gram-positive and gram-negative bacteria, including strains that have already developed resistance.27 Rather than replacing current antibiotics, these therapies would restore or amplify their activity, potentially extending the useful life span of established treatments. Ideally, such agents would combine broad applicability with practical advantages in manufacturability, cost, safety, and clinical deployment.
Many of the emerging modalities discussed earlier, including phage therapy, host-directed therapies, antibody-based antibacterial strategies, microbiome-directed treatments, and polymeric antibiotics, could contribute to this type of pathogen-general strategy. By either enhancing host defenses, targeting bacterial virulence mechanisms, reshaping microbial ecosystems, or sensitizing bacteria to existing drugs, these approaches may complement traditional antibiotics and expand the range of tools available for managing resistant infections.
Antimicrobial resistance will not be solved by simply discovering more versions of the same antibiotics that bacteria have already evolved resistance to. Instead, progress will likely come from a broader therapeutic ecosystem that combines multiple complementary strategies. Phage therapies, antibody-based antibacterial agents, microbiome-directed treatments, host-directed approaches, and engineered antimicrobial polymers each operate through fundamentally different mechanisms that can either bypass traditional resistance pathways or restore the effectiveness of existing drugs. As these modalities mature and enter clinical development, they may work alongside conventional antibiotics rather than replace them outright. Expanding the antimicrobial toolkit in this way could provide the flexibility needed to address the diverse and evolving challenges posed by multidrug-resistant pathogens.




